内耳异位再生毛细胞的ADF/destrin发育表达及其作用机制
2020-12-24涂澄宇张亮彭佳南昌大学第四附属医院耳鼻喉科江西南昌330001
涂澄宇 张亮 彭佳 (南昌大学第四附属医院耳鼻喉科,江西 南昌 330001)
哺乳动物的内耳毛细胞比较脆弱,极易遭受噪声、毒素等因素的影响,导致其毛细胞受到损害,而这些受损的毛细胞无法自发再生,这也是它们后天出现获得性耳聋的主要原因。destrin会通过化学当量促使纤维形肌动蛋白(F-actin)迅速实现解聚。现阶段,由哺乳动物各类组织内纯化得来的destrin,它会与F-actin原聚体相互结合,并把actin分子拆下来,并依托剪切、聚集到与之对应的F-actin〔1〕。新生或成年鼠内耳依然有部分细胞具备前体或者干细胞特征,上述细胞支持在体外培养条件下形成细胞球,并能够分化为毛细胞样细胞。与光感受器相比较,听觉和前庭感觉上皮相似,出现更为复杂的感觉区,在以上感觉区主要任务在于合理平衡声音、感受刺激,并通过化学信号转化而来的毛细胞〔2〕。本文探究胚胎14.5 d直至其出生后4 d听觉、半规管壶腹上皮发育中的ADF/destrin表达。
1 材料与方法
1.1材料与试剂 青链霉素、谷氨酰胺均由Gibco公司提供;山羊、驴封闭血清均从MILLIPORE公司购入; 60 mm和100 mm培养皿、冻存管等产品均由Corning公司提供; 35 mm培养皿由Falcon公司提供;驴抗小鼠-Rho二抗(1∶200)、山羊抗小鼠cy5二抗(1∶200);山羊抗兔-488二抗(1∶200);ADF/destrin(兔来源单克隆抗体)(1∶100~1∶200)、E-cadherin(小鼠来源单克隆抗体)、Western印迹试剂盒(南京建成生物科技有限公司)、PCR试剂盒(大连TakaRa)等。
1.2孕鼠天数计算 挑选C57BL/6型生长健康的小鼠,雌、雄鼠数量依次为6只、2只,并于实验的上午10点将3只雌鼠、1只雄鼠放到同一个窝内,24 h后分开以后,次日上午除掉胚胎计作E0.5,依次类推从同窝后第15天、19天分别计作E14.5、E18.5。实际实验中,同窝后第15天、19天下午8点、6点所取的胚胎依次计作E14.5、E18.5。
1.3胚胎小鼠实施耳蜗处理 采用颈部脱臼把孕鼠处死后,通过剖腹方法快速将子宫取出,将其置于1×磷酸盐缓冲液(PBS,pH7.4)溶液内,并对胚囊进行相应的剥离,把得到的胚胎放在直立荧光纤维镜下把胚胎放入冰冷的4%多聚甲醛溶液内,并解剖内耳,将其固定到4%多聚甲醛溶液内,并在4℃冰箱内保存。
1.4耳蜗全基底膜铺片 在显微镜下把固定完成的耳内移到1×PBS(pH7.4)溶液内实现解剖操作,将耳蜗基底膜完整的取出来,除掉表面盖膜等,并将其放在0.1%TritonX-100的1×PBS溶液内室温下进行20 min漂洗处理,并在48培养孔板中开展免疫染色处理。
1.5组织离体实施病毒培养 采用脱臼的方法将新生小鼠实施处死处理,并采用75%酒精完成消毒,并把颞骨听泡部位取出并盛到经过灭菌处理的玻璃皿内,在镜下将耳蜗基底膜取出来。由35 mm×10 mm培养皿中事先加入已经涂抹0.1%多聚莱氨酸盖玻片,并增加1.0 ml的血清培养液中,移动基底膜保证匍匐贴壁。在37℃温箱条件下实施孵育过夜。次日,把其换为无血清培养液,转入Ad5-EGFP-math1或Ad5-EGFP病毒,促使其终浓度成为病毒(PFU)1.0×108。在此基础上,根据组织贴壁细胞生长状况每隔1 d、2 d更换1次培养液。在37℃环境的温箱进行孵育,CO2浓度为5%,湿度控制为95%,并在转入病毒后6 d、12 d取出培养皿观察。
1.6开展扫描处理 利用Zeiss LSM510mets激光共聚焦的显微镜展开处理,依次设置不同的波长,分别为488 nm(FitC)、633 nm(cy5)、543 nm(rhodamin)的激光,对其进行40倍放大操作,从耳蜗基底膜自顶层慢慢向下完成扫描,设定层厚数值为0.5 μm,图像分辨率设定为2 048×2 048。
1.7小鼠内耳毛细胞体外诱导分化体系建立及ADF/destrin敲除体获得 取出实验小鼠的内耳耳蜗,将其Cochlear组织小心放入35 mm的平皿中,加入适量无菌PBS缓冲溶液;在24孔板中,小心加入无菌的圆形小玻片,其直径约为14 mm,并在小玻片上加入一定量的尾胶稀释液,置于37℃中孵育15 min;接着,将内耳组织小心放入含有尾胶的24孔板中,再继续加入1 ml的DMEM/F12培养基;轻轻的平稳的将24孔板小心置于培养箱中,整个过程保证耳蜗不动,紧密贴着鼠尾胶;小鼠cochlear培养过程中,可以加入适量100 mg/L 4-OH Tamoxifen诱导Cre活性;继续培养7~10 d后,观察并拍照。
1.8小鼠内耳毛细胞体外诱导分化体系建立 本研究参考刘亚青等〔3〕研究方法,成功地构建了小鼠内耳毛细胞体外诱导体系,其内耳支持细胞来源于Math1-GFP小鼠。ADF/destrin敲除体获得是通过与生物公司合作,由生物公司上海斯莱克实验动物有限公司构建。
1.9切片制作 先修整小鼠内耳组织,经流水冲洗30 min后,取掉多余固定液;接着依次将其浸泡在70%、80%、90%和100%的乙醇中各脱水1 h,在100%的乙醇中多脱水1 h;接着,在二甲苯中,使得脑组织透明化,处理2次,分别为15 min和10 min;最后,将其依次放入恒温箱中,融化石蜡,每次1 h,重复3次。再将处理后的内耳组织按照顺序包埋成块状,连续冠切片,每张切片保证其厚度约为5 um,切片完成后,用于后续组织形态学和免疫组化学检测。小鼠内耳组织HE染色方法如下:将上述准备好的切片,浸入二甲苯中,2次,每次约为10 min;浸入100%乙醇中,2次,每次约为10 min;再依次浸入90%、80%、70%酒精及蒸馏水中,各约5 min;再用配制好的苏木素染色液染色8 min,染色完成后,用自来水冲洗;分化20 s(由70%乙醇加1%盐酸配制而成)后,浸入蒸馏水中处理5 min;接着按照70%、80%乙醇,浸泡5 min;再浸入含有90%伊红醇溶液中,浸泡10 min;再浸入100%乙醇中,2次,每次5 min;再浸入二甲苯中,2次,每次5 min;最后,在中性树胶中封片,显微镜下观察。
1.10统计学处理 采用SPSS20.0软件行t检验。
2 结 果
2.1正常小鼠耳蜗基底膜中ADF/destrin表达 胚胎期14.5 d(E14.5)至出生后的第4天(P4)期间,ADF/destrin处于耳蜗中分布状况。在E14.5状态下,destrin 处于分散分布的状况,且毛细胞和支持细胞均有与之对应的表达,如图1所示。在出生后第1天(P1)时,destrin大多出现在毛细胞纤毛及其内指细胞中;P4时主要存在至此细胞质内有表达。
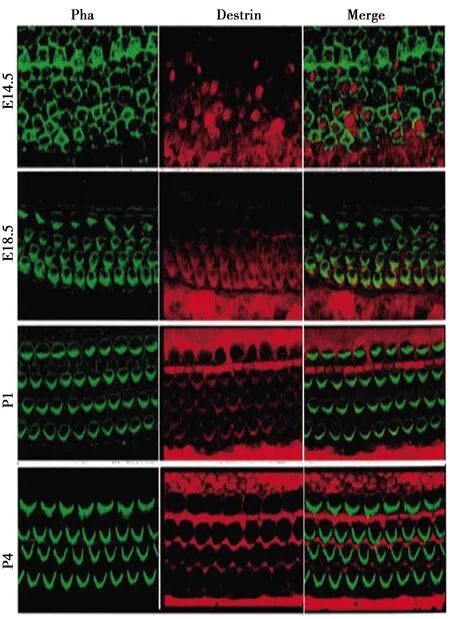
图1 小鼠从胚胎至出生后各阶段耳蜗中ADF/destrin表达(×200)
2.2dADF/destrin在小鼠内耳毛细胞体外诱导分化体系中的表达 随着诱导时间的增加,Math1的表达量也在增加,同时,dADF/destrin的表达也在增加,并且在第6天达到高峰期。见表1,图2。

表1 相关基因表达分析

图2 Western 印迹检测培养不同时间Math1、dADF/destrin表达水平
2.3敲除dADF/destrin抑制小鼠毛细胞发育 通过Cre/Loxp系统,成功地构建了小鼠dADF/destrin敲除体,当小鼠在E13~E15时,通过灌胃的方式,给予小鼠Tamoxifen,进一步诱导Cre活性。同时,当胎鼠为20 d时,通过组化法分析突变胎鼠的内耳表型。与正常小鼠相比,dADF/destrin敲除小鼠体积更小,发育迟缓,且内耳结构也有一定的絮乱。见图3、图4。
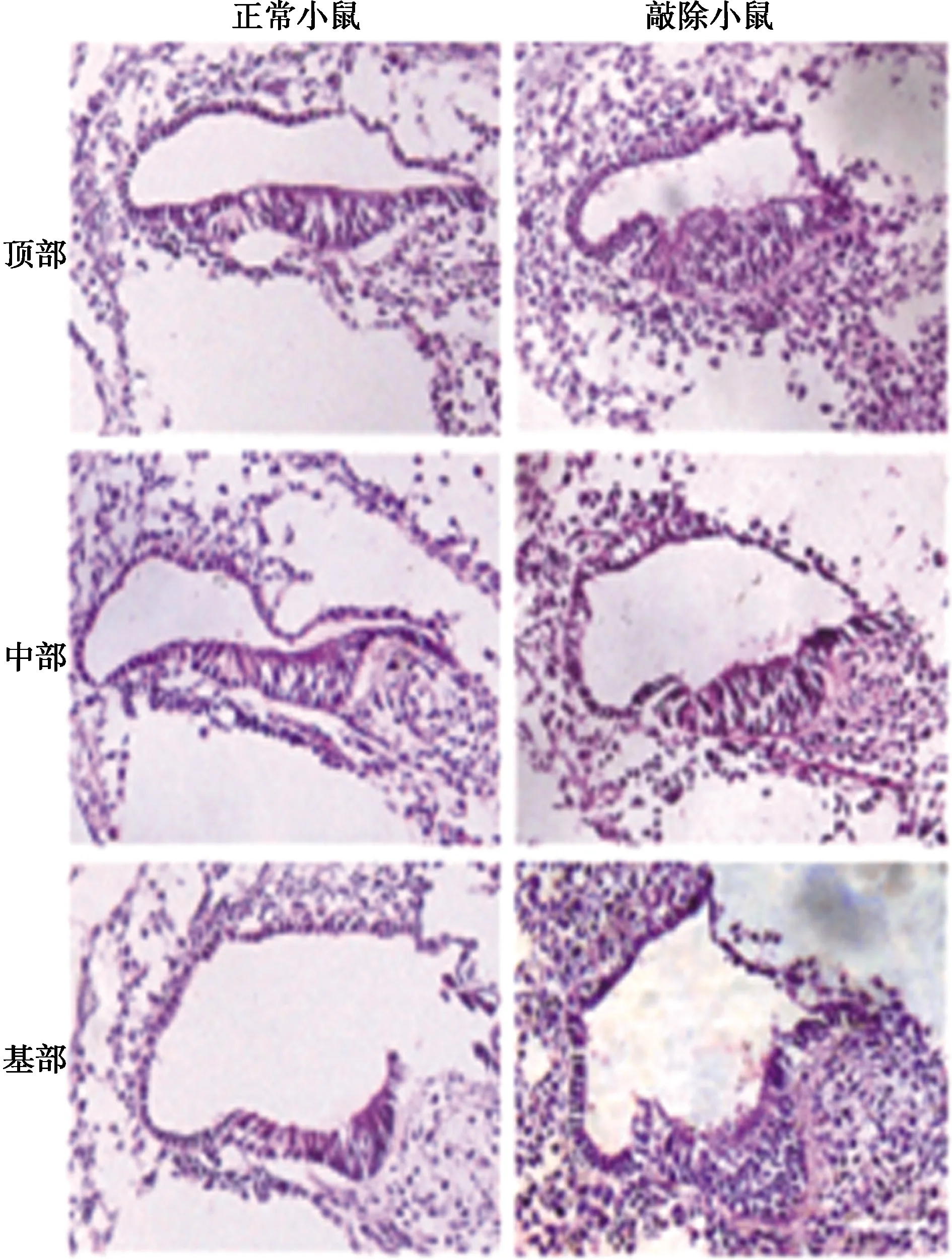
图3 HE染色分析正常小鼠与敲除小鼠内耳结构(×200)
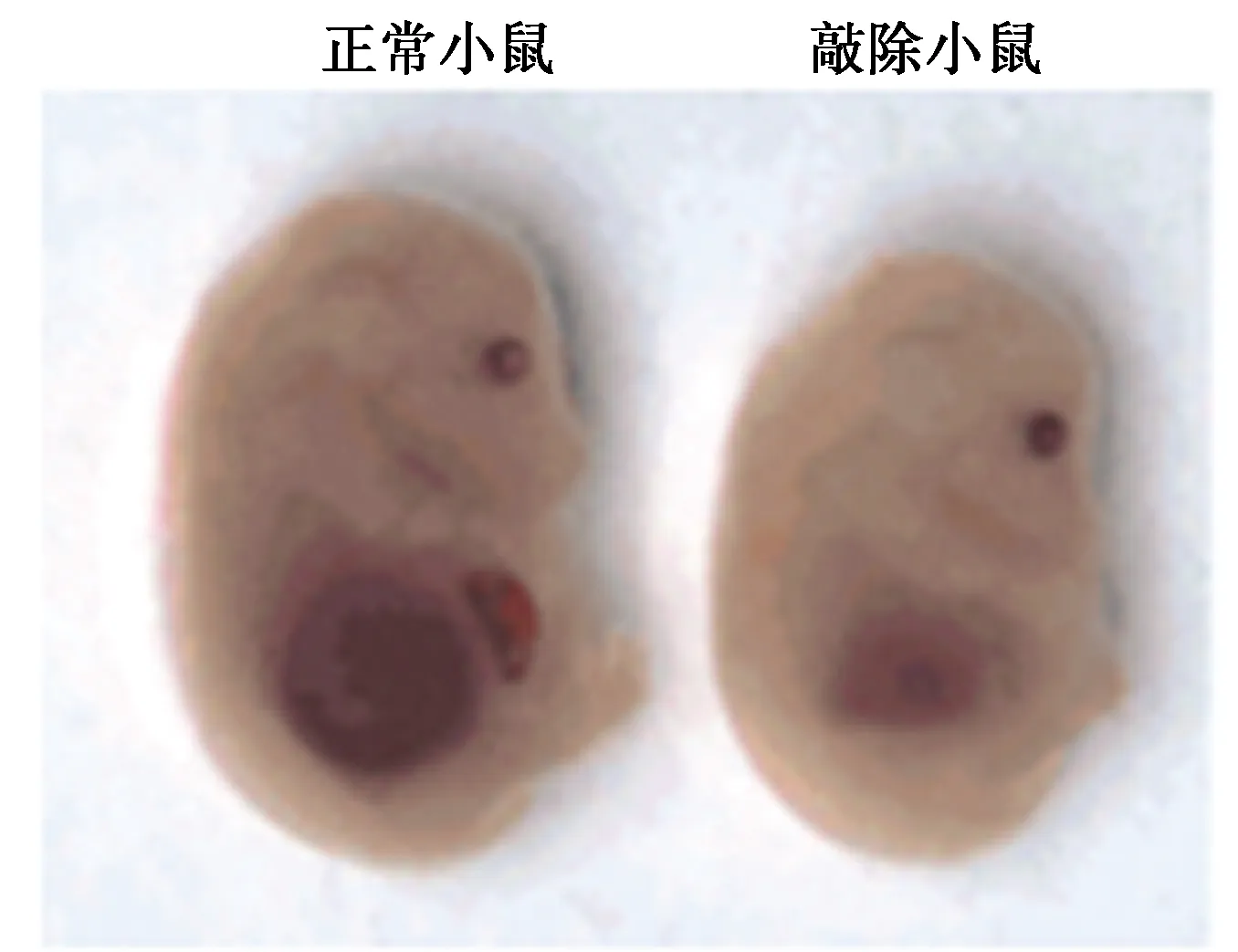
图4 正常小鼠与敲除小鼠胚胎发育
3 讨 论
哺乳动物内耳形态发生改变属于比较比较复杂的过程,毛细胞是处在内耳的机械感受器细胞,可以把声音、运动信号等转换为电化学信号。哺乳动物的耳蜗毛细胞再生能力有一定的限制,利用药物或基因干预可以对其实施恰当的修复。根据小鼠实验结果可知,细胞周期蛋白大部分使用激酶抑制剂(CDKI)p27等,确保哺乳动物支持细胞可以顺利进入细胞周期作为重要的调节因素〔4〕。为保证哺乳动物的毛细胞受到相应的损伤后可以及时修复,以上蛋白均可当做短暂性基因将靶标敲除。基于此,通过逆转录病毒Atoh1因子保障成熟的细胞可以转化为毛细胞。除此之外,异位再生毛细胞发育时期是否出现平面细胞极性(PCP)改变受到多数研究者的重视和关注。ADF/destrin属于肌动蛋白解聚因子中不可缺少的一部分,其他成员包含cofilin2、cofilon1。ADF/destrin出现在海马锥体细胞全部的兴奋性突触,且突出前、后结构均有出现,这为动物ADF/destrin相关问题研究提供一定的参考〔5〕。但ADF敲除突变体发生突触前保留其生理功能之外,ADF失活并没有影响动物的神经元及其他功能师傅恢复正常。通过研究野生型果蝇发育情况可知,视网膜细胞通过5倍的延长,tsr RS突变体因伸展阶段缺失,导致黏附连接变宽〔6〕。
感觉细胞、成熟神经元大部分源自细胞表面,也称为原纤毛细胞突起。原纤毛基体并没有得到生产,在参与细胞分裂过程中有丝分裂纺锤体内,如果原纤毛发生有丝分裂前会受到再次吸收,这一细胞周期和纤毛形成密切的联系,说明原纤毛发育环节发生细胞增殖及分化产生重要的影响。原纤毛具体功能展现在细胞天线上,其具体表现为及时接收细胞外信号,用于合理调节动物大脑发育状况〔7,8〕。在上述研究背景下,原纤毛发育与出现疾病相关性成为研究的热点问题,细胞表面突起多数参与机械刺激中,其主要缺点会使得部分器官出现功能性疾病。原纤毛作为化学及机械信号两方面进行处理的感受器,它不单能够整合细胞周围信息,各整合操作发挥着重要的作用〔9〕。从某些分裂细胞视角分析,原纤毛对细胞是否要重新进入其周期并保持静止状态产生影响,使原纤毛发育一个关键的事件在于前体细胞/干细胞之间的不对称分裂,进而发生相同或展现出不同命运的子代。有丝分裂过程中分裂面成为判断肝细胞分裂是否对称具有重要的影响,原纤毛极易对分裂后2个子代细胞有影响。随着医学界对原纤毛问题研究的更深入,能够获取关于感觉发育机制的新见解〔3,10〕。ADF失活并不会影响神经元分化及其突触功能。本研究结果表明,耳蜗听觉上皮及其前庭感觉上皮发育阶段ADF/destrin出生4 d后主要出现在支持细胞中,这一情况与毛细胞、支持细胞负责的各功能有着紧密的联系;本研究提示,这一时期动纤毛上肌动蛋白解聚活动非常强,之所以发生这种状态,可能于动物的纤毛发育或退化情况有着密切的联系。本研究结果还表明,ADF/destrin在椭圆囊、半规管壶腹毛细胞表皮板到小鼠出生后仍然有所表达,表明与前庭感觉上皮相比较,耳蜗听觉发育更早。
