耳蜗动纤毛在听觉系统中的作用研究进展
2023-07-31章天成洪国栋付小龙柴人杰
章天成,洪国栋,李 贺,付小龙,柴人杰
(1.温州医科大学,浙江 温州 325035;2.山东第一医科大学·山东省医学科学院,山东 济南 250117;3. 温州医科大学附属第一医院,浙江 温州 325000;4.东南大学,江苏 南京 211189)
听觉是人类最重要的感觉之一,它的发生和发展与人类的进化密不可分。人类的日常沟通交流离不开听觉,遗传性的缺陷或者耳毒性药物的滥用等都会造成听觉系统不可逆的损伤,给人们的正常生活和工作造成极大的困难。在毛细胞上还有一种由微管蛋白组成的特殊的初级纤毛—动纤毛,近几年科学家们发现动纤毛结构异常会导致毛细胞上的纤毛束极性紊乱,纤毛束的方向不再一致,从而导致严重的听力损失。本文综述动纤毛在耳蜗中的发育过程及其在听觉发生过程中的作用,为预防由动纤毛异常导致的听力损失提供理论参考。
1 听觉系统与毛细胞纤毛结构
外周听觉系统主要由外耳、中耳和内耳组成。其中内耳不仅要负责声音信号的传导过程,还要具备将外界声音的机械能信号转换为神经电信号的功能。耳蜗中复杂声音信号转换的关键部位称为柯蒂氏器,柯蒂氏器是基底膜中负责听觉产生的精妙结构,其中包含有多种重要细胞类型,如毛细胞和支持细胞,毛细胞又可分为一排内毛细胞和三排外毛细胞,毛细胞和支持细胞以相互嵌入的方式排列,在外毛细胞的顶表面有三排特化后的呈现“V”形状或者“W”形状的由肌动蛋白组成的静纤毛束,三排静纤毛束以阶梯状排列,靠近内部的一排纤毛较短,最外一排纤毛最长[1~3],见图1[4]。外毛细胞上的静纤毛束统一朝向非神经轴侧,这一特性称之为毛细胞的平面细胞极性(planar cell polarity,PCP),这些静纤毛束插入盖膜当中,当声波信号传入内耳后会引发盖膜和纤毛束之间的剪切运动,进而打开或关闭纤毛上特有的机械电转导通道,这是听觉发生过程中声音信号转换的关键过程,而内毛细胞上的纤毛束主要呈现出“一”字形或者“U”形,似乎并不参与机械电转导过程[5~11]。
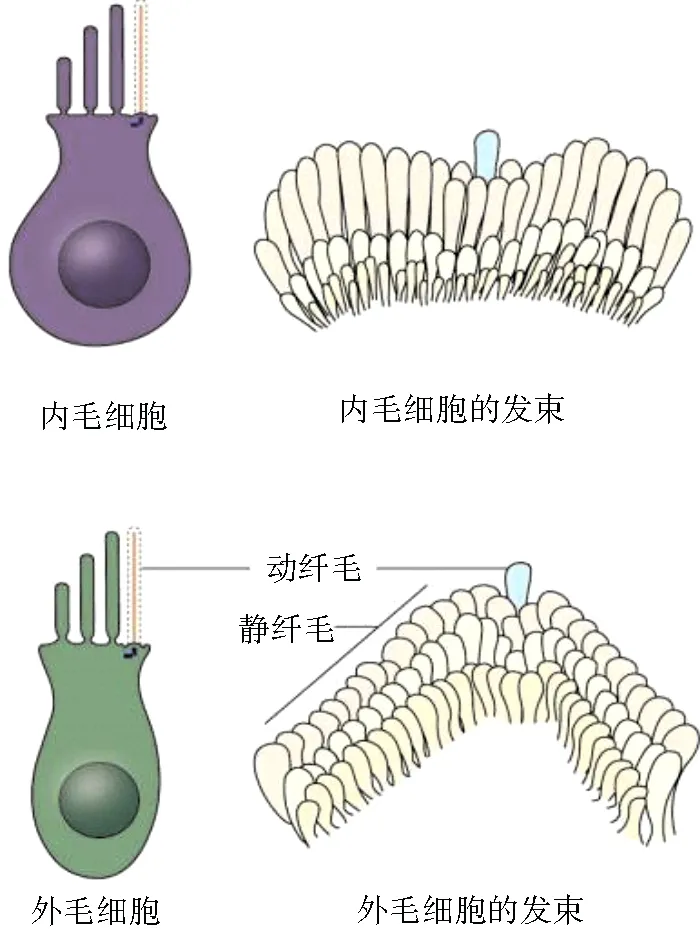
图1 耳蜗毛细胞和纤毛束的示意图
2 纤毛束在听觉发生过程中的作用
听觉发生过程是一个复杂且精细的过程,它需要多个听觉器官与大脑听觉中枢协同作用,且缺一不可,若听觉发生过程中的某个环节异常则会引起不同程度的听力损失。耳廓首先将外界声音收集,然后经外耳道、鼓膜传入中耳,随后再经过听小骨、卵圆窗膜等部位将声波的振动信号传入内耳,再由内耳将声波信号转换为听觉神经冲动,耳蜗内的螺旋神经元将这些神经冲动传递到听觉中枢中的各个核团,最终由听觉皮层对声音信号进行分析处理并产生听觉[12~14]。耳蜗是听觉系统中的重要感受器官,耳蜗结构或功能受到损伤后将会导致严重的听力损失。声音信号转导过程的关键步骤是毛细胞上静纤毛束的有效偏转,当声音的机械能信号导致静纤毛束发生偏转后会打开静纤毛束上的机械电转导通道,随后的离子交换过程使毛细胞兴奋并产生声音的电信号,因此,纤毛束的结构和功能的稳态维持在听觉发生过程中是极为重要的[15]。
3 动纤毛的结构及发育
3.1 动纤毛的结构纤毛是一种位于细胞表面的突起结构,广泛分布于几乎所有真核细胞表面[16]。纤毛的结构主要包括基体、轴丝、纤毛膜和纤毛基部。纤毛基体是由母中心体发育而来,主要由微管蛋白组成。纤毛膜是细胞膜的延伸,但其成分跟普通细胞膜有较大的差异。轴丝是由9条环形排列的双联体微管蛋白束及其附属蛋白构成,轴丝从纤毛基部开始组装、延伸。部分纤毛微管中间存在一对中心微管,根据纤毛是否有中心微管及可否自主运动可以将纤毛分为“ 9+2”运动型、“ 9+2 ”不动型、“ 9+0 ”运动型和“ 9+0 ”不动型四种类型[17]。哺乳动物内耳毛细胞的动纤毛是一类特殊的纤毛,属于“ 9+2 ”不动型[18]。虽然它有外侧动力臂和径向辐条,但没有内侧动力臂,见图2[4]。因此,尽管外侧动力臂使动纤毛保留了一些运动功能,但由于缺乏内侧动力臂,它只能被动摆动,而不能自主运动[19]。
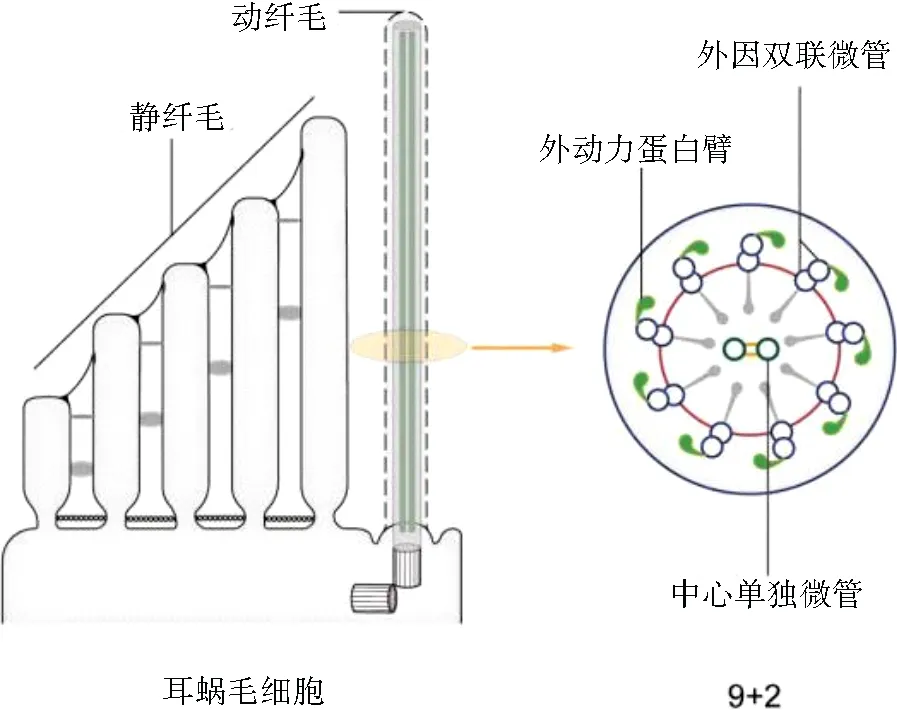
图2 动纤毛及其横截面的模型
3.2 动纤毛的发育听纤毛发育是一个受到严格调控的过程[20]。哺乳动物发育早期阶段,每一个毛细胞的顶表面都覆盖着以肌动蛋白为骨架的微绒毛。在小鼠胚胎期的第15天,动纤毛在耳蜗毛细胞顶表面的中心开始发育[21],周围被非常短的微绒毛围绕,并从毛细胞顶表面的中心逐渐向非神经轴侧迁移,起到了引导作用,协调了富含肌动蛋白的新生静纤毛束的形态发生和定向[22]。所以,静纤毛束的定向被认为是由动纤毛引导的[23, 24]。与毛细胞顶表面上的静纤毛不同的是,动纤毛不直接介导听觉发生中的机械电转导过程,但部分保留了负责毛细胞对声音刺激反应的运动特性。随后,耳蜗毛细胞中动纤毛在小鼠出生后的第八天开始退化,P14天动纤毛完全消失[25]。
4 动纤毛在听觉系统中的作用
4.1 动纤毛在毛细胞成熟过程中的作用在毛细胞发育早期,动纤毛从毛细胞顶表面的中心逐渐向非神经轴侧迁移的同时引导了静纤毛束的形成与定向。以鸡的耳蜗为例来分析动纤毛在毛细胞发育过程中可能的作用,毛细胞纤毛束在成熟过程中大致可以分为四个阶段,在胚胎第8天(E8)之前为第一阶段,毛细胞前体细胞经历了终末分裂,毛细胞开始分化;第二阶段(E8~ E13),毛细胞上的动纤毛向纤毛束的非神经轴侧逐渐移动,与动纤毛相邻的静纤毛增长,同时和动纤毛之间形成特殊的动纤毛连接,随后三排静纤毛形成特定的阶梯结构;第三阶段(E13~ E16),静纤毛的延长过程中止,静纤毛开始变宽,其底部逐渐变细,表皮板开始形成;第四阶段(E16 ~P3),静纤毛继续伸长,直到静纤毛成熟为止[20],以上的描述共同提示了动纤毛在耳蜗毛细胞发育成熟过程中扮演着十分重要的角色。随后,越来越多的证据显示动纤毛在毛细胞发育及功能的调控过程中发挥重要作用。例如,双皮层蛋白结构域蛋白2a(doublecortin domain containing 2a,DCDC2a)表达于感觉毛细胞的动纤毛和支持细胞的初级纤毛中,有研究利用基因编辑技术敲除大鼠DCDC2a基因后,敲除鼠的毛细胞动纤毛结构被破坏,静纤毛束形态改变及长度显著增加,进而引起了严重的不可逆性听力损失[26]。Bardet-Biedl综合征(Bardet-Biedl syndrome,BBS)临床表现为耳聋、智力发育迟缓、先天性肥胖及视网膜营养不良等,当小鼠Bbs1、Bbs4等BBS综合征基因发生突变后会导致耳蜗动纤毛与静纤毛的连接异常,进而导致静纤毛束形态及其极性发生改变[27, 28]。
4.2 动纤毛通过调节PCP来调控毛细胞的发育在小鼠耳蜗发育早期,毛细胞上具有一根动纤毛和很多的微绒毛,大部分微绒毛会随着年龄的增长而慢慢退化,最终只留下三排特化后的微绒毛形成“V”形的纤毛束,也被称为静纤毛。在纤毛束特化这一过程中动纤毛也会逐渐退化直至完全消失,最终只留下动纤毛残迹,因此,动纤毛被广泛认为不直接参与听觉过程。目前,关于耳蜗动纤毛的报道主要是其参与纤毛束PCP的调控。所有毛细胞的静纤毛束朝向一致,形成了毛细胞的平面细胞极性[29]。毛细胞PCP的形成是一个受到精细调控的过程[22, 24],进一步的研究显示动纤毛通过调控毛细胞PCP从而在毛细胞发育及功能中发挥重要作用。例如,PCP核心蛋白编码基因的缺陷,如Ltap/Vangl2, Dvl1/2, Fz3/6以及Celsr1,都显示不同程度的动纤毛紊乱[30~35]。在PCP缺陷的基因敲除小鼠中,非对称分布的PCP膜相关蛋白定位紊乱,导致耳蜗汇聚伸展和纤毛束极性的异常,从而导致了严重的听力障碍[26]。也有研究报道发现Dyx1c1基因缺失会导致原发性纤毛运动障碍,纤毛的运动能力丧失,此外Dyx1c1敲除小鼠耳蜗毛细胞上动纤毛的结构以及退化过程被破坏了,同时还发现伴有暂时性的毛细胞PCP方面的缺陷[36, 37]。有研究指出驱动蛋白家族成员3A(Kinesin family member 3A,Kif3A)通过纤毛和非纤毛机制调节毛细胞极化,内耳中Kif3A的缺陷导致动纤毛的缺失和PCP紊乱类似的表型(即耳蜗导管的缩短和发束的扁平形态),最终导致听觉障碍[38]。
5 总结与展望
耳蜗毛细胞上的动纤毛是基于微管蛋白组成的具有经典“9 + 2”结构的纤毛,然而与常见的运动纤毛不同的是耳蜗动纤毛不具备运动能力,又因为耳蜗动纤毛在听力形成之前就会完全退化而只留下一个残迹,所以研究人员普遍认为动纤毛对听觉的发生不是必要的。目前的研究主要集中于动纤毛结构破坏和纤毛束PCP之间的相关性上,然而仍然没有直接的证据表明动纤毛结构改变是导致PCP改变的直接原因,比如,某种基因缺陷在破坏动纤毛结构的同时也会直接影响纤毛束极性的改变,这种情况不能说明动纤毛结构改变是导致PCP紊乱的直接原因。因此,详细阐明遗传缺陷导致动纤毛或纤毛束PCP变化的分子机制有利于我们进一步了解动纤毛和PCP之间的关联性,同时也能完善动纤毛在耳蜗中扮演的其他可能存在的角色。总之,遗传缺陷型耳蜗毛细胞PCP的紊乱是由于动纤毛结构异常间接导致的还是由于遗传缺陷直接导致的仍有待更深入的研究。
