亚热带生态系统5种修复树种的光合特征1)
2024-05-11袁媛徐克芹曾平生姚甲宝孙洪刚欧阳琼肖斌
袁媛 徐克芹 曾平生 姚甲宝 孙洪刚 欧阳琼 肖斌
(中国林业科学研究院亚热带林业实验中心,江西·分宜,336600) (中国林业科学研究院亚热带林业研究所) (中国林业科学研究院亚热带林业实验中心)
当前我国林草事业进入以生态保护修复为主的发展阶段,主要任务是保护管理林草资源,推进大规模国土绿化和生态修复保护,提高陆地自然生态系统质量和稳定性。我国部分生态公益林因人为干扰或种质低劣、经营管理不当等原因,形成林下植被单一、水土流失严重、生态服务功能低下的经营型低效林。按照我国颁布的GB/T 18337.1—2001《生态公益林建设导则》,允许进行以提高森林生态系统稳定性或改善生态功能为主要目标的森林抚育、更新采伐等森林经营活动。采取小块状或团状采伐形成林窗,促进植物更新,是调整森林树种组成、改善森林结构和提升森林功能的重要推动力[1]。苏宇等[2]对柏木低效林开窗补植银木的研究发现,阔叶树种的引入能有效促进森林土壤养分循环过程,加速退化林分修复。因此,在经营过程中,通过透光抚育疏伐、开窗补植乡土树种和林下植被更新等改造措施,能有效改善低效林结构和组成的空间异质性,加快群落演替进程,最终实现系统稳定性和生态服务功能的提升。
光合作用是植物生长和物质积累的基础,影响植物个体的“存活-生长”,在一定程度上反映了植物对环境的适应能力[3]。受低效林改造所形成的林窗大小、形状、所处地理环境以及边缘木状况等因素的影响,不同属性林窗内光环境以及由光照所导致的热量、水分和营养条件等资源分布也会发生相应的变化,影响更新幼苗的增补、定植与分布[4-6],因此低效林开窗补植树种应首先考虑引入树种幼苗的光合特征和对林窗环境适应能力。苦槠(Castanopsissclerophylla)、木荷(Schimasuperba)、浙江楠(Phoebechekiangensis)、刨花楠(Machiluspauhoi)和红楠(Machilusthunbergii)是南方地区造林和生态修复常用的地带性常绿阔叶树种,通过对5种树种的生长特性[7-9]、生理生态[10-14]、化学计量[15-17]和栽培示范[18-19]较为深入的研究,但对5种生态修复常用造林树种生物学特性认识还不足,尤其对其光合特性方面的比较研究甚少。因此研究这些树种的光合特性对其在生态修复中的应用和栽培模式的选择具有重要意义。
本研究以苦槠、木荷、浙江楠、刨花楠和红楠2年生容器苗为试验材料,在测定其生长指标、叶绿素质量分数、光合和叶绿素荧光参数的基础上,分析不同树种的光合特性差异,探究不同树种幼苗光合能力的差异,确定树种在低效公益林改造中的配置,为低效公益林质量提升提供理论依据。
1 试验区概况
试验在中国林业科学研究院亚热带林业实验中心苗圃内进行,试验区地处江西省新余市分宜县,境内为低山丘陵地貌,以红壤为主,土层深厚,土壤肥沃。气候属亚热带湿润性气候,雨水充沛、光照充足、气候温和、无霜期长,年均温17.2 ℃,年降水量1 600 mm,全年无霜期达270 d。试验区自然资源丰富,森林覆盖率达65.76%。乔木树种主要有杉木(Cunninghamialanceolata(Lamb.) Hook.)、马尾松(PinusmassonianaLamb.)、湿地松(PinuselliottiEngelm)和毛竹(Phyllostachysheterocycla(Carr.) Mitford cv.Pubescens)等,林下植被由檵木(Loropetalumchinense(R. Br.) Oliv.)、柃木(EuryajaponicaThunb)、乌饭(VacciniumbracteatumThunb)、杜鹃(RhododendronsimsiiPlanch.)等灌木以及蕨类、芒萁(Dicranopterisdichotoma(Thunb. ) Berhn.)、芭茅(Miscanthusfloridulus(Labill.) Warb)等组成。
2 材料与方法
2.1 试验材料
试验盆栽苗放置在苗圃中心位置,四周放置其他普通容器苗,以确保处理环境无差异。以苦槠、木荷、浙江楠、刨花楠和红楠为试验材料,选用规格为15 cm×20 cm的无纺布袋为栽植容器,无纺布袋中装入等量同种配制基质,有机质质量分数为6.45 g·kg-1,全氮和全磷质量分数分别为0.35、0.36 g·kg-1,水解氮、有效磷和速效钾质量分数分别为46.23、10.47、92.5 μg·g-1。于2021年1月15日播种,5月中旬选择生长状况良好的芽苗移栽至无纺布袋,保持通风、湿润及适当光照,按正常生长苗木管理两年。
2.2 光合特性参数测定
光响应曲线测定:于2022年7—8月份晴朗天气,使用Li-6400便携式光合仪(LI-COR Inc, Nebraska, USA),在每天09:00—11:30和15:00—17:00进行测定。设定测前诱导光强(PAR)为1 000 μmol·m-2·s-1,温度为(26±1)℃,CO2摩尔分数为400 μmol·mol-1,空气流速为0.5 L·min-1,叶室中光合有效辐射分别为2 000、1 500、1 000、600、300、200、100、80、50、20、0 μmol·m-2·s-1,每一光强下停留200 s,同一树种重复6次。
光合指标测定:选取同生长状况基本一致种试验树种3株,每株选择顶芽下第3~4枚完全展开的3枚成熟叶片,使用Li-6400便携式光合仪(LI-COR Inc,Nebraska,USA)进行测定。设定测前温度为(26±1)℃,CO2摩尔分数为400 μmol·mol-1,空气流速为0.5 L·min-1,光合有效辐射(PAR)为1 000 μmol·m-2·s-1,稳定后记录净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2摩尔分数(Ci)。光合水分利用效率(RWUE),RWUE=Pn/Tr。
叶绿素荧光参数测定:先将叶片置于暗处进行暗反应,20 min后,使用Li-6400便携式光合仪(LI-COR Inc,Nebraska,USA)测定暗反应下的初始荧光(Fo)、最大荧光(Fm)、可变荧光(Fv)、光系统Ⅱ(PSⅡ)潜在光化学活性(Fv/Fo)和光系统Ⅱ(PSⅡ)最大光化学效率(Fv/Fm)。
2.3 叶绿素质量分数的测定
8月下旬,选取同种试验树种3株长势相似的健康苗木,在苗木中部相同位置,取成熟叶片洗净擦干,去除中脉剪碎,用电子顶载天平(0.01 g)称取0.2 g至具塞离心管,用80%丙酮定容至10 mL,密封后置于暗处静置72 h。测定时取上清液于比色皿中,以80%丙酮作为对照,用分光光度计测定叶绿素提取液在波长为663和645 nm下的吸光值[20-22]。
2.4 各树种生长指标及生物量测定
同年12月测定各树种生长指标及生物量。每个树种选取6株生长良好、长势相近的苗木,用游标卡尺测量地径,钢卷尺测量苗高,电子顶载天平测量植株鲜生物量,将其置于烘箱70 ℃烘至恒质量,测定植株干物质质量和植株含水率。
2.5 数据处理
采用Microsoft Excel 2010对基础数据进行整理;以叶子飘等[23]的光合计算软件4.1.1为处理平台,拟合试验树种光响应曲线,并进一步估算出表观量子效率(α)、最大净光合速率(Pn,max)、光饱和点(LSP)、光补偿点(LCP)和暗呼吸速率(Rd)等特征参数;以Origin 2017为作图软件绘制光响应曲线、光合指标和叶绿素荧光指标图;利用SPSS20.0进行数据处理分析,采用单因素方差分析和最小显著差异法(LSD)对数据进行差异性比较,采用Pearson相关性分析对数据进行相关性检验。
光响应曲线表达方程为:
式中:Pn为净光合速率;α为表观量子效率;β、γ为修正系数;I为光合有效辐射;Rd为暗呼吸速率。
光饱和点(LSP)计算公式:
式中:LSP为光饱和点;β、γ为修正系数。
最大净光合速率(Pn,max)计算公式:
式中:Pn,max为最大净光合速率;α为表观量子效率;β、γ为修正系数;Rd为暗呼吸速率。
3 结果与分析
3.1 供试树种的生长指标
由表1可知,各树种幼苗地径差异显著,苦槠地径最大,木荷次之,浙江楠、刨花楠和红楠的地径显著低于苦槠和木荷。苗高38.7~48.7 cm,其中,苦槠苗高显著高于浙江楠、刨花楠和红楠,木荷次之,且与其他树种无显著差异。供试树种的鲜物质质量存在显著差异,苦槠鲜物质质量显著高于刨花楠和红楠,木荷鲜物质质量和浙江楠与其他树种无显著差异。苦槠干物质质量显著高于刨花楠和红楠,木荷和浙江楠干物质质量次之,且与其他树种无显著性差异,刨花楠和红楠的干物质质量较小。各植株含水率位50%~60%,其中刨花楠含水率最最高,显著高于其他树种,5种树种苗木的含水率由高到低顺序为刨花楠、浙江楠、苦槠、木荷、红楠。
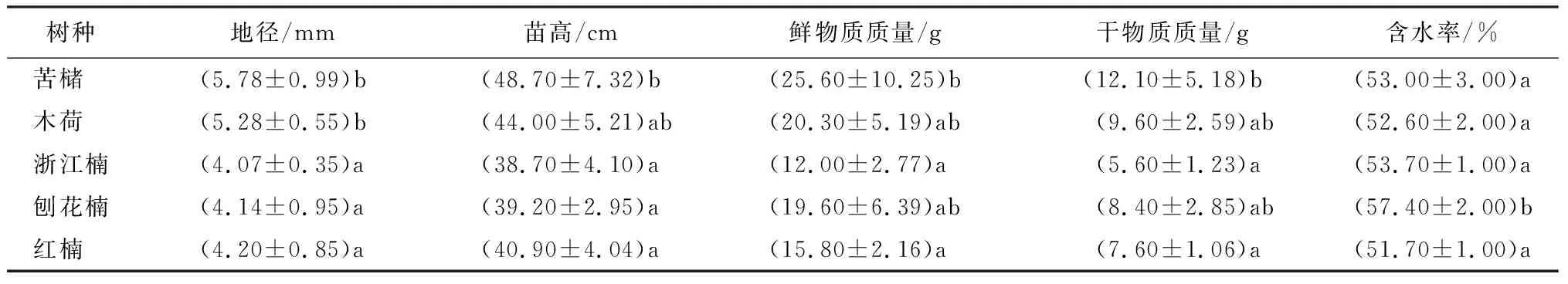
表1 供试树种的生长指标
3.2 供试树种的叶绿素质量分数
由表2可知,苗木叶片叶绿素a质量分数在供试树种间存在显著差异,其中,红楠叶绿素a质量分数最低且显著低于苦槠、木荷和浙江楠,刨花楠次之,且与其他树种无显著差异,木荷叶绿素a质量分数为最高;各树种叶绿素b质量分数为0.10~0.53 mg·g-1,且各树种间差异不显著,叶绿素b质量分数由大到小顺序为浙江楠、木荷、苦槠、刨花楠、红楠;各树种间的总叶绿素质量分数存在显著差异,木荷和浙江楠的总叶绿素质量分数较高,且显著高于红楠,苦槠和刨花楠次之,且与其他树种间差异不显著,红楠的总叶绿素质量分数为0.37 mg·g-1,为供试树种的最小值。
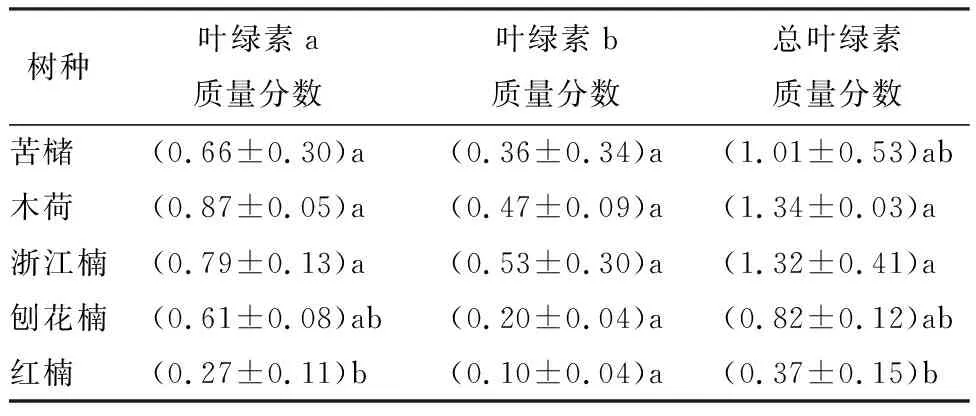
表2 供试树种的叶绿素质量分数 mg·g-1
3.3 供试树种光响应曲线及特征参数
由图1可知,供试树种的光响应曲线变化规律较为一致,大致可分为3个阶段,第一阶段随着光照强度的增强,净光合速率呈直线上升趋势;第二阶段随光照强度的持续增强,净光合速率缓慢增加;第三阶段光照强度达到植物光饱和点时,净光合速率随光照强度增强而有所下降。当光合有效辐射小于600 μmol·m-2·s-1时,供试树种的净光合速率均随光合辐射增加显著升高,且各树种间存在显著差异;当光合有效辐射大于600 μmol·m-2·s-1时,各树种净光合速率随光合辐射增加逐渐趋于光饱和点,随后出现光抑制现象。不同树种达到光饱和点的光合辐射差异显著,浙江楠、刨花楠和红楠在光合辐射为600 μmol·m-2·s-1左右时达到光饱和点,木荷和苦槠在光合辐射大于1 000 μmol·m-2·s-1时逐渐达到光饱和点,表明浙江楠、刨花楠和红楠对弱光利用能力好,在弱光环境下适应性更强,而木荷和苦槠需要更高的光照强度才能到达最大净光合速率,更加适宜在强光环境下生长。
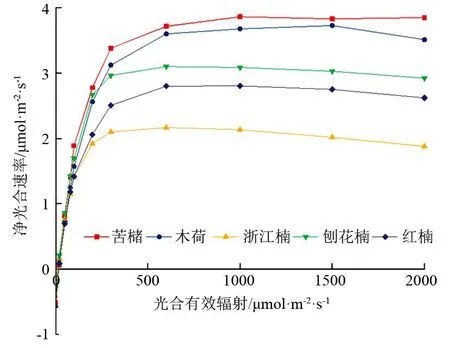
图1 各供试树种的光响应曲线
由表3可知,供试树种的表观量子效应值为0.030~0.048,且树种间差异显著(P<0.05),其中红楠的表观量子效应值显著大于其他树种,表明红楠对弱光的利用效率最高。各树种最大净光合率差异显著,其中,浙江楠的最大净光合率值最小,显著小于苦槠、木荷和刨花楠,苦槠的最大净光合率值最大,显著高于浙江楠、刨花楠和红楠,最大净光合率由大到小排列顺序为苦槠、木荷、刨花楠、红楠、浙江楠。不同树种对光环境适应情况差异较大,苦槠的光饱和点值和光补偿点值均高,表明强光环境下适应力较强;刨花楠的光饱和点值和光补偿点值均低,为耐阴性植物,在弱光环境下适应性更强,木荷的光饱和点值较高且光补偿点值较低,可利用的光能范围较广,对光环境具有较强的适应性。供试树种的暗呼吸速率值存在显著差异,红楠的暗呼吸速率值最大,对光能产物的消耗最大;苦槠次之,且与其他树种无显著差异;木荷、浙江楠和刨花楠的暗呼吸速率值较小,且显著低于红楠;暗呼吸速率由大到小的顺序为红楠、苦槠、浙江楠、刨花楠、木荷。
3.4 各供试树种光合指标的差异
由表4可知,5种树种的净光合速率(Pn)存在显著差异,苦槠的Pn值最大且显著高于浙江楠、刨花楠和红楠,浙江楠的Pn值显著低于苦槠和木荷,木荷、刨花楠和红楠间的Pn值无显著性差异。苦槠的气孔导度(Gs)值最大,且显著高于浙江楠、刨花楠和红楠;木荷气孔导度次之,与其他树种无显著差异;浙江楠、刨花楠和红楠的Gs值较小,且树种间没有显著性差异。各供试树种间的胞间CO2摩尔分数(Ci)值差异显著,苦槠和浙江楠的Ci值显著高于木荷和红楠;刨花楠的Ci值显著高于木荷;木荷和红楠Ci值较低,且各树种间无显著性差异。供试树种的蒸腾速率(Tr)值为0.625~1.567 mmol·m-2·s-1,木荷的Tr值最大且显著高于其他树种,Tr值由大到小的顺序为木荷、苦槠、刨花楠、红楠、浙江楠。各树种的光合水分利用效率(RWUE)为2.709~3.898 μmol·mmol-1,且各树种间无显著差异,RWUE值由大至小顺序依次为苦槠、红楠、刨花楠、浙江楠、木荷。
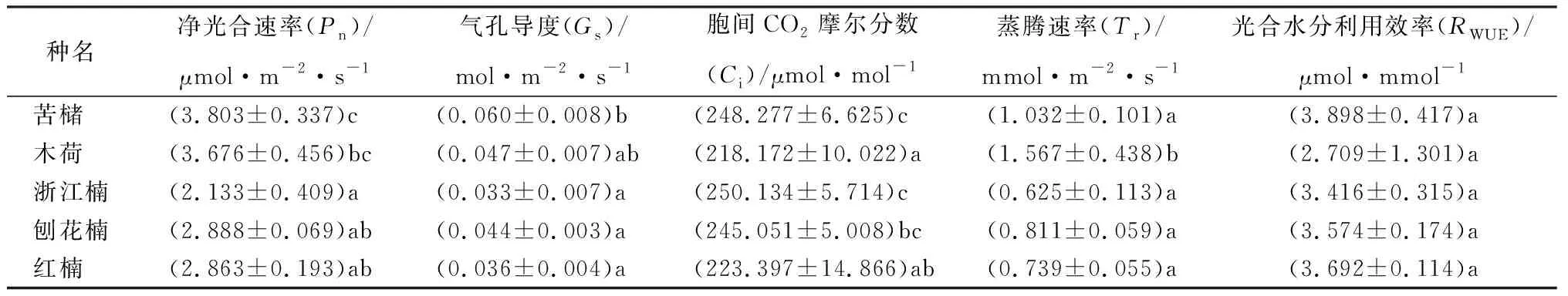
表4 各供试树种的光合指标
3.5 各树种叶绿素的荧光参数
由表5可知,初始荧光(Fo)和最大荧光(Fm)分别为光系统Ⅱ(PSⅡ)反应中心处于完全开放和完全关闭时的荧光产量,供试树种的Fo和Fm均没有显著性差异,表明5种树种幼苗的叶绿素质量分数和电子传递活性相近。Fv/Fo反映光系统(PS)Ⅱ的潜在荧光活性,Fv/Fm为光系统(PS)Ⅱ最大光化学效率,在暗处理条件下,光系统PSⅡ的初始光能转化效率;供试树种的Fv/Fo值和Fv/Fm值由大到小顺序均表现为苦槠、红楠、木荷、刨花楠、浙江楠,表明苦槠的PSⅡ反应中心的潜在荧光活性和光能转化效率均为供试树种最高。

表5 各供试树种的叶绿素荧光指标
3.6 各影响因子的相关性
由表6可知,供试树种的地径与苗高、最大净光合速率(Pn,max)、光饱和点呈极显著正相关(P<0.01);苗高与Pn,max呈极显著正相关(P<0.01),与光饱和点、光补偿点、Fv/Fo和Fv/Fm显著正相关(P<0.05);Pn,max和光饱和点、Fv/Fo、Fv/Fm显著正相关(P<0.05);光饱和点与Fv/Fo、Fv/Fm显著相关(P<0.05);Fv/Fo与Fv/Fm之间极显著正相关(P<0.01)。
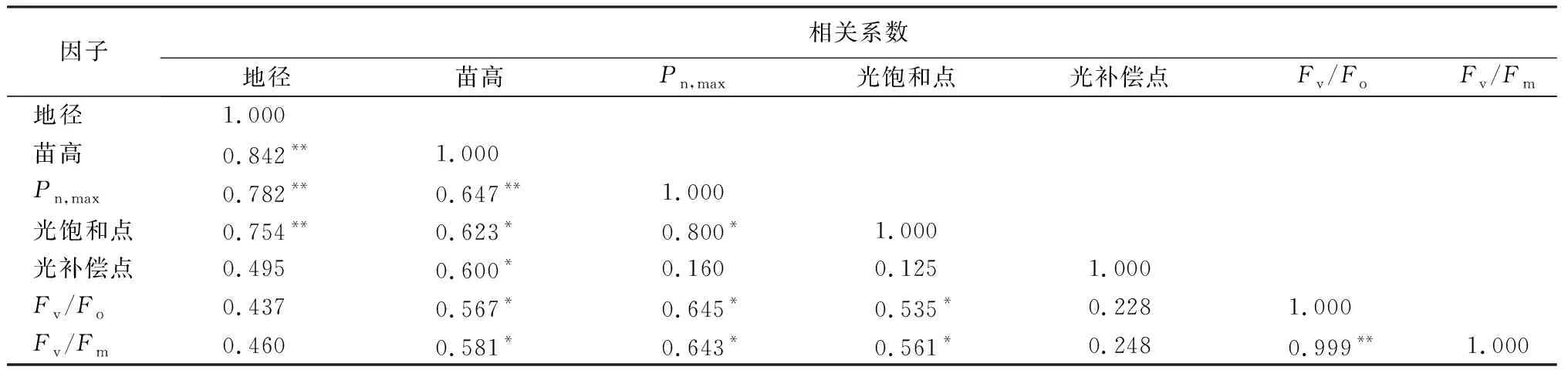
表6 各影响因子相关系数
4 结论与讨论
地径和苗高是反映幼苗生长状况的最直观的指标。对生长特征参数、光响应曲线特征参数和叶绿素荧光参数进行相关性分析发现:地径与最大净光合速率呈极显著相关且相关性系数最大,表明树种光合能力越强,植株的生长状况越好;地径与光饱和点极显著正相关,苗高与光饱和点、光补偿点显著正相关,说明供试树种幼苗生长还与其光环境适应能力密切相关;苗高还与光系统(PS)Ⅱ潜在荧光活性(Fv/Fo)和光系统(PS)Ⅱ最大光化学效率(Fv/Fm)显著相关,叶绿素荧光参数也是影响苗高的重要因素。因此,光合特性可以作为预测供试树种幼苗生长状况的重要指标。
光合作用是植物物质积累的主要来源,与植物生长发育密切相关,主要受自身遗传特性影响,常用光响应曲线分析植物光合作用过程[24-25]。植物净光合速率随光合有效辐射变化主要呈现两种趋势,一种是随光照强度增加,净光合速率持续增加并在达到最高点后保持不变,一种是随光照强度增加,净光合速率增加至光饱和点后出现光抑制现象[26-28]。本研究中,供试树种的光响应曲线均呈现第二种变化趋势,表现为随光照强度增加,净光合速率先直线上升后缓慢增高至最高点再有所下降的变化趋势,但不同树种达到最大净光合速率的光照强度(光合有效辐射)有所差异。浙江楠和刨花楠的最大净光合速率在光照强度(光合有效辐射)为600 μmol·m-2·s-1,木荷和苦槠在光照强度(光合有效辐射)超过1 000 μmol·m-2·s-1净光合速率达到最高点,表明浙江楠和刨花楠为耐阴性植物,适宜在弱光环境下栽植,木荷和苦槠对强光利用能力更强,适宜栽植在光照强度高的环境。供试树种均出现光抑制情况,说明光照过强会减缓5种树种幼苗的生长,适宜栽植在有一定遮阴条件的林窗环境。
植物光合作用是受自身和环境共同影响的复杂过程,光合指标是植物光合作用的生理特征及其环境适应性的具体体现,各指标之间相互制约和促进[29]。本研究中,5种树种的Pn、Gs和Ci之间存在显著性差异,而Tr和RWUE在树种间差异不显著。其中,供试树种的Pn和Gs呈现相同的变化趋势,Pn和Gs由大到小的顺序为苦槠、木荷、刨花楠、红楠、浙江楠,表明气孔开闭程度是影响供试树种光合速率的主要因素,这与黄娟等[30]的研究较为一致。叶绿素荧光指标是揭示环境对植物光合作用影响的内在探针,反映了植物内部对光能的吸收、转化及光合生理状况[31-32]。参试树种间的Fo和Fm差异不显著,而Fv/Fo和Fv/Fm在树种之间差异显著。光系统(PS)Ⅱ潜在荧光活性(Fv/Fo)、光系统(PS)Ⅱ最大光化学效率(Fv/Fm)与最大净光合速率、光饱和点呈显著正相关,说明叶绿素荧光参数能在一定程度上反映植物光合性能,光系统(PS)Ⅱ反应中心的潜在活性越高、光能转化效率越高,植物光合能力越强。
光合特征参数反映了植物光合作用的生理特性及环境适应性。表观量子效率反映植物在弱光条件下的光能利用效率,正常生长环境下的实测值一般为0.03~0.05[33],5种树种表观量子效率值范围在0.030~0.048,表明供试树种幼苗没有受到光抑制。通常,植物的表观量子效率越高和光补偿点越低,植物耐阴性越好。本研究中,供试树种的表观量子效率和光补偿点呈相反的变化趋势,与前人研究结果相似[34]。5种供试树种中,红楠的表观量子效率高、光补偿点低,表明其对弱光的利用能力较好;苦槠的表观量子效率较低、光补偿点较高,更适宜在强光环境下生长。光饱和点和光补偿点是表征植物对光环境适应性的指标,光饱和点越高,强光利用能力越强,光补偿点越低,弱光利用能力越强。本研究发现,苦槠阳性特征显著,木荷的光环境适应性较强,浙江楠、刨花楠和红楠对弱光的利用能力较好。
研究和比较不同树种间光合特性和生长能力,对于评价其对环境的适应能力具有重要意义[18]。根据树种光合特性的差异筛选和配置树种,可为低效公益林改造提供科学依据[35]。综上所述,5种亚热带生态修复树种在苗木生长、叶绿素含量和光合特性之间差异显著,在进行低效公益林改造林窗补植时应结合树种光合特性进行选择。苦槠光饱和点、光补偿点最高,表观量子效率最小,对强光环境具有较好的适应性,在林窗补植时可选择林窗中心位置进行栽植。木荷光饱和点高且光补偿点较低,光环境适应能力强,既能较好地适应强光环境又有较强的弱光利用能力,适合配置在林窗近中心位置。浙江楠光饱和点、光补偿点、暗呼吸速率较小且表观量子效率较大,对光能的利用性较好,尤其是对弱光利用能力较强,适宜栽植在林窗边缘和中林窗位置。刨花楠最大净光合速率较高,光补偿点、光饱和点、暗呼吸速率较低,有较高的光合潜力,具有较好的耐阴性,但对强光利用能力较弱,可将其作为阴坡林窗和小林窗的栽植树种。红楠表观量子效率最大,光饱和点和光补偿点较小,对弱光的利用能力强,耐阴性较好,可选择林窗边缘位置或者阴坡、半阴坡环境栽培。
