高剂量葡萄糖对小鼠肌管细胞自噬表达的影响
2019-06-03罗胜男苏震黄佩佩陈如意左一丹胡菲菲
罗胜男,苏震,黄佩佩,陈如意,左一丹,胡菲菲
(温州医科大学附属第一医院 肾内科,浙江 温州 325015)
糖尿病患者持续高血糖状态可引起骨骼肌萎缩,称为糖尿病肌病[1],临床表现为肌肉量减少,肌肉无力和身体机能下降[2]。蛋白质代谢紊乱导致肌 肉质量和功能的下降是糖尿病管理中最具挑战性的问题之一。胰岛素可抑制蛋白分解并促进蛋白合成,调节机体血糖水平。1型糖尿病出现骨骼肌萎缩主要由胰岛素缺乏引起,而2型糖尿病则是因为骨骼肌对胰岛素敏感性下降,泛素-蛋白酶系统和自噬-溶酶体系统活性增高,蛋白质代谢失衡[3]。自噬是介导细胞内功能失调的细胞器和蛋白质降解的重要分解代谢机制[4],当自噬被过度活化,蛋白质被过度降解,肌纤维功能受损,最终导致骨骼肌萎缩。高剂量葡萄糖对骨骼肌细胞自噬稳态的影响尚不清楚,本研究主要探索高剂量葡萄糖对骨骼肌自噬的影响,同时通过动态监测自噬流,进一步比较高剂量葡萄糖作用不同时间自噬表达差异。
1 材料和方法
1.1 材料
1.1.1 C2C12细胞株及慢病毒:小鼠C2C12成肌细胞株购自中国科学院细胞库,货号GNM26,使用含10%胎牛血清、100 U/mL青霉素和100 μg/mL链霉素的DMEM培养液(美国Gibco公司)培养,置于37 ℃,5% CO2恒温孵育箱中。串联荧光标记LC3(mRFP-GFPLC3)慢病毒购自上海吉凯基因有限公司。
1.1.2 试剂:Western blot一抗包括:抗LC3B购自美国Abcam公司,抗Beclin-1购自美国Cell Signaling Technology公司,抗体稀释度为1:2 000和1:1 000。抗β-actin购自美国Cell Signaling Technology公司,抗体稀释度为1:1 000。PCR引物包括:LC3上游引物:5’-GCCTGTCCTGGATAAGACCAA-3’和下游引物:5’-ACCATGCTGTGCTGGTTGAC-3’。Beclin-1 上游引物:5’-AACTCGCCAGGATGGTGTCT-3’和下游引物:5’-CTTGAGTCTCCGGCTGAGGT-3’。
1.2 方法
1.2.1 C2C12细胞培养、诱导分化和实验处理: 当C2C12成肌细胞融合达70%~80%,更换为含2%马血清的分化培养基培养2 d,诱导成肌细胞分化成肌管细胞。然后分3组,分别予正常剂量(5.5 mmol/L) 和高剂量(17 mmol/L、30 mmol/L)葡萄糖浓度的培养基培养,以5.5 mmol/L组作为正常对照组。之后每2 天更换相应葡萄糖浓度的培养基。6、12、24、36、48、60和72 h后分别收集细胞,提取蛋白和RNA,检测自噬表达情况。
1.2.2 慢病毒转染:根据说明书使用串联荧光标记LC3(mRFP-GFP-LC3)慢病毒转染C2C12成肌细胞,感染复数(MOI)为100,转染助剂Polybrene(购自上海吉凯基因有限公司)终浓度为5 μg/mL,转染8 h后更换培养基为生长培养基,再培养72 h后进行后续实验(参考方法1.2.1中的实验处理)。使用4%多聚甲醛将单层肌管细胞固定于盖玻片上,并用DAPI染料(购自北京索莱宝科技有限公司)进行细胞核染色,将盖玻片倒扣于载玻片上,封片后用于激光共聚焦显微镜的观察。
1.2.3 慢病毒原理:串联荧光标记LC3(mRFP-GFPLC3)慢病毒结构中,GFP信号对pH的敏感性高于RFP,在溶酶体酸性环境下发生淬灭[5]。因此,合并图像中GFP和RFP荧光信号的共定位,呈现黄色荧光斑点(即GFP+/RFP+-LC3)表示自噬体。相反,只有RFP荧光信号(即GFP-/RFP+-LC3)呈现红色,表示自噬溶酶体。
1.2.4 Western blot检测:收集肌管细胞,使用含有1 mmol/L苯甲基磺酰氟(PMSF)的RIPA裂解缓冲液(购自上海碧云天生物技术有限公司)冰上裂解细胞,提取细胞蛋白,使用BCA蛋白浓度检测试剂盒(购自上海碧云天生物技术有限公司)测量总蛋白量。随后行SDS-PAGE电泳分离蛋白,并电转至PVDF膜上,5%脱脂奶粉封闭2 h,一抗4 ℃孵育过夜。用TBST洗膜4次,每次10 min,然后用山羊抗兔二抗(购自美国Biosharp公司)孵育1 h,抗体稀释度1:2 000。再次用TBST洗膜4次,每次10 min,采用增强的化学发光试剂(购自美国Thermo公司)进行检测,用自动扫描仪分析蛋白印迹并使用图像分析软件数字化定量分析条带灰度值。
1.2.5 实时荧光定量PCR:使用TRIzol试剂(购自美国Invitrogen公司)提取总RNA,并采用反转录试剂盒(购自美国Thermo公司)合成cDNA。使用SYBR Premix Ex Taq II(购自日本Takara公司)在CFX Connect实时PCR检测系统(购自美国Bio-Rad公司)上进行定量PCR。每个样品均设置3个复孔,并以GAPDH作内参进行校正。
1.3 统计学处理方法 采用SPSS21.0软件进行统计学分析。计量资料用±s表示,多组间比较用单因素方差分析,组间两两比较用LSD-t法进行分析;培养时间与葡萄糖浓度的交互作用采用双因素方差分析。P<0.05为差异有统计学意义。
2 结果
2.1 3组不同剂量葡萄糖作用不同时间肌管细胞C2C12自噬相关蛋白和基因表达结果 3组不同葡萄糖浓度的培养基分别培养6和12 h,自噬标记蛋白LC3和Beclin-1表达水平组间差异无统计学意义(P> 0.05),见图1。与5.5 mmol/L组比,30 mmmol/L 组培养24 h,LC3-II和Beclin-1蛋白水平显著降低,并且LC3的mRNA水平也显著降低,差异有统计学意义(P<0.05),见图2A。当不同剂量的葡萄糖培养基培养36 h,30 mmol/L组LC3、Beclin-1蛋白和基因表达较5.5 mmol/L组升高,其中,LC3-II蛋白水平较5.5 mmol/L组明显升高,差异有统计学意义(P<0.05),见图2B,这与培养时间24 h的结果相反。30 mmol/L组培养48 h后LC3-II和Beclin-1 蛋白水平较5.5 mmol/L组均显著增加,差异有统计学意义(P<0.05),见图3A。PCR结果与Western blot结果一致,与5.5 mmol/L组相比,30 mmol/L组LC3和Beclin-1 mRNA水平均明显升高,差异有统计学意义(P<0.05),见图3A。30 mmol/L葡萄糖浓度的培养基培养C2C12肌管细胞60和72 h后结果与48 h一致,自噬活性显著高于5.5 mmol/L组,见图3B-C。不同时间点LC3-II和Beclin-1 蛋白表达量变化规律一致,见图4。培养时间与葡萄糖浓度存在交互作用,差异有统计学意义(P<0.001),高剂量葡萄糖30 mmol/L组培养72 h,自噬蛋白表达量最高。

图1 6 h和12 h 3组C2C12肌管细胞LC3-II和Beclin-1蛋白表达结果(n=6)

图2 24和36 h 3组C2C12肌管细胞LC3-II和Beclin-1蛋白和mRNA表达结果(n=6)
2.2 高剂量葡萄糖作用不同时间C2C12细胞串联荧光标记LC3表达结果 在C2C12肌管细胞的激光共聚焦显微镜图像中,存在一些RFP和GFP荧光斑点,并且许多荧光斑点在合并图像中共定位,表明存在自噬体和自噬溶酶体。随葡萄糖浓度的升高,合并图像中不仅黄色斑点数量明显增加,RFP荧光斑点数量也显著增加,差异有统计学意义(P<0.05),见图5。与5.5 mmol/L组比,高剂量葡萄糖30 mmol/L 组培养60 h和72 h后RFP和GFP荧光斑点数量均明显升高,差异有统计学意义(P<0.05),见图6-7。
3 讨论
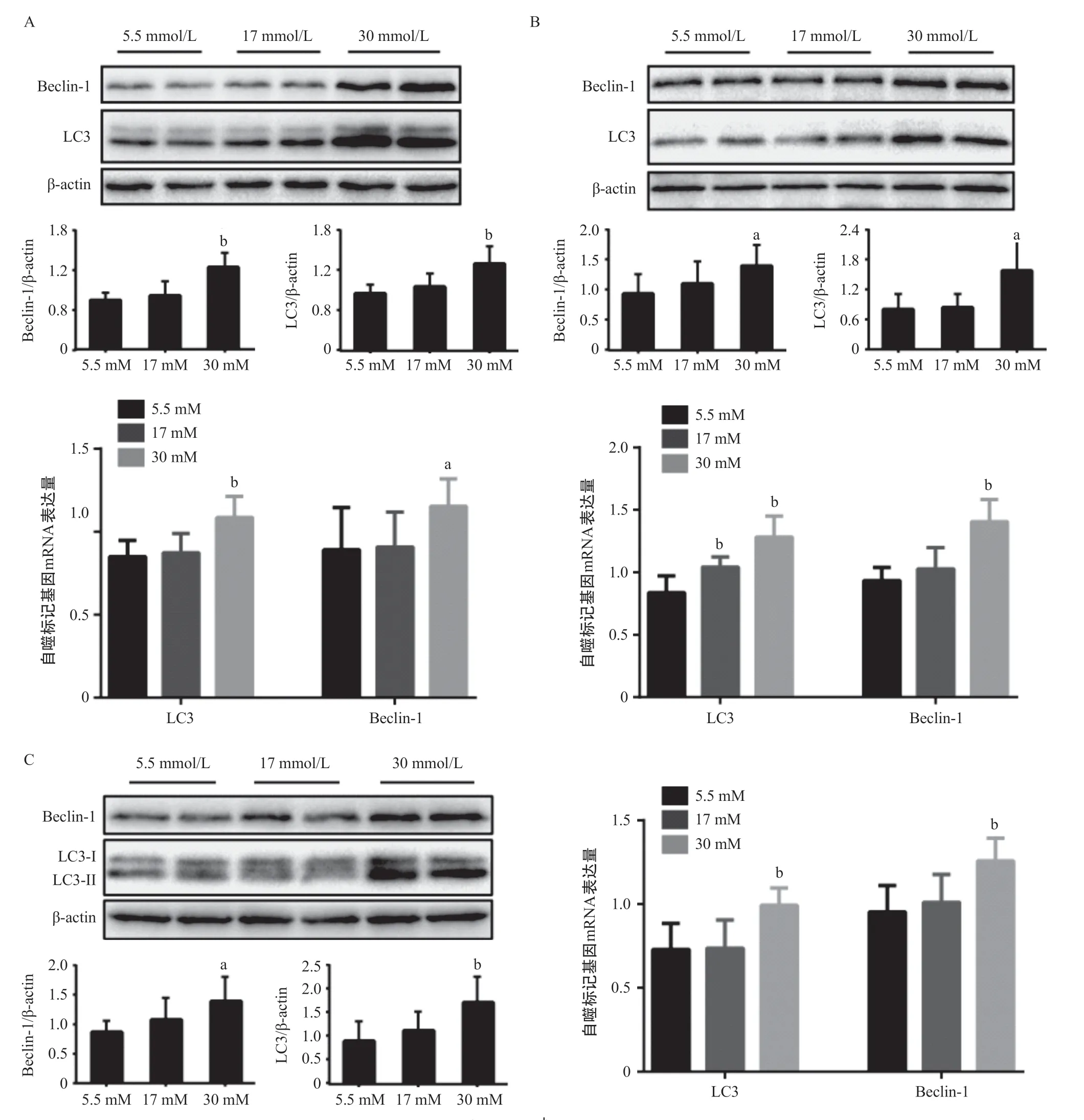
图3 48、60和72 h 3组C2C12肌管细胞LC3-II和Beclin-1蛋白和mRNA表达结果(n=6)
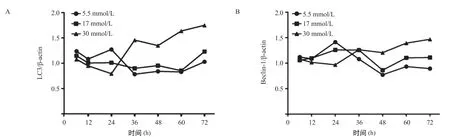
图4 不同时间点自噬标记蛋白表达变化趋势线图(n=6)

图5 48 h激光共聚焦显微镜检测LC3荧光光斑(×400,n=12)
1型和2型糖尿病均可影响骨骼肌正常代谢,导致肌肉质量和力量下降,称糖尿病肌病[6]。与糖尿病其他并发症(如血管病、视网膜病、肾病和神经病)相比,糖尿病肌病经常被忽视且尚未被临床诊断。基于骨骼肌在体内葡萄糖代谢中的关键作用,糖尿病肌病还会加重其他并发症的疾病进展。但糖尿病肌病的发生机制尚不清楚,有研究指出糖尿病会引起骨骼肌细胞的兴奋-收缩耦联机制障碍[7],高剂量葡萄糖培养的肌纤维细胞表现为横管形态肿胀,单一电场刺激引起动作电位诱导的Ca2+释放增加。高剂量葡萄糖诱导的横管结构的破坏和Ca2+释放改变可能在糖尿病肌病肌无力的发病机制中起到主要作用。此外,胰岛素抵抗可影响蛋白质代谢,机体高血糖和慢性炎症状态致细胞出现氧化应激、细胞器损伤以及自噬过度活化[8],加速机体蛋白质的消耗[9]。
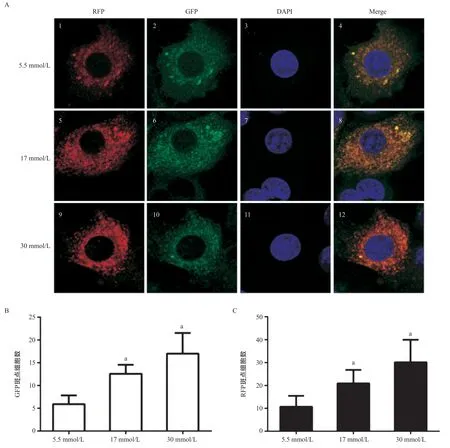
图6 60 h激光共聚焦显微镜检测LC3荧光光斑(×400,n=12)

图7 72 h激光共聚焦显微镜检测LC3荧光光斑(×400,n=12)
自噬-溶酶体系统是细胞内主要的蛋白质降解系统,对于细胞更新和体内代谢平衡至关重要[10],在维持肌肉质量和肌纤维稳态方面也发挥着重要作用[11-12]。在骨骼肌中,自噬囊泡将衰老的细胞器和错误折叠的蛋白质转运至溶酶体,并与溶酶体融合形成自噬溶酶体,然后囊泡内的物质被降解产生氨基酸、核苷酸、葡萄糖和脂肪酸用于细胞物质合 成[11,13]。近年来,自噬-溶酶体系统过度活化已被证实可引起蛋白质代谢紊乱,导致骨骼肌萎缩[14]。某些疾病状态下引起的骨骼肌萎缩,包括饥饿、衰老[15]、脓毒血症[16]、慢性肾功能不全[17]和癌症[18]等,均与自噬过度活化有关。营养缺乏自噬因子-1(nutrient deficiency autophagy factor-1,Naf-1) 是自噬调节因子,在Naf-1基因敲除小鼠中,骨骼肌结构和性能因自噬增强而显著受损[19]。因此,自噬稳态是保持肌肉质量和功能所必需的[20]。
当自噬微管相关蛋白1 轻链3(LC3-I)转化为 LC3-II并融入自噬泡时,就提示形成了成熟的自 噬[21]。同时,Beclin-1的募集是自噬起始的信号,可以调节自噬上游[22]。因此,LC3-II和Beclin-1 蛋白表达水平与自噬体的数量成比例。本研究使用这2种自噬蛋白和自噬标记基因来评估细胞外高剂量葡萄糖对C2C12肌管细胞自噬的影响,结果表明细胞外高剂量葡萄糖可破坏肌管细胞自噬稳态。另外,还发现不同的培养时间具有不同的自噬通量。早期阶段,当高剂量葡萄糖培养C2C12肌管细胞24 h,与 5.5 mmol/L组比,自噬标记基因和蛋白表达均被显著抑制,这与之前的报道[23]一致。3种葡萄糖浓度的培养基分别培养肌管细胞48、60和72 h,Western bolt和PCR结果分析均显示高剂量葡萄糖30 mmol/L 组自噬体合成显著增加。
自噬是一个动态多步骤的过程,自噬体的积累可能表明自噬生成增加或自噬体清除减少。自噬标记物如LC3-II和Beclin-1 的分析,只能近似估计自噬通量。为进一步探讨细胞外高剂量葡萄糖对骨骼肌细胞自噬通量的影响,本研究采用了串联荧光标记LC3(mRFP-GFP-LC3)慢病毒稳定转染C2C12成肌细胞,动态监测自噬体成熟过程[5]。肌管细胞RFP和GFP荧光斑点的定量分析显示,高剂量葡萄糖培养48、60和72 h后,自噬体和自噬溶酶体均显著增加。结果表明,长时间高浓度葡萄糖环境下培养C2C12肌管细胞可诱导自噬过度活化,且高剂量葡萄糖对肌管细胞自噬的作用与培养时间相关。
本研究在不同时间评估高血糖对骨骼肌细胞自噬作用的重要性,从而推测,糖尿病患者骨骼肌萎缩的触发事件可能是自噬-溶酶体系统被过度激活、机体蛋白质降解增加、长期高血糖状态最终导致糖尿病肌病。
