新疆传统发酵食品中乳酸菌的分离鉴定及其生长代谢特性
2017-04-26凌空翟磊姚粟宋振杨玉新程池
凌空,翟磊,,姚粟,宋振,杨玉新,程池*
1(中国食品发酵工业研究院,北京,100015) 2(新疆中亚食品研发中心(有限公司),新疆 乌鲁木齐,830001)
新疆传统发酵食品中乳酸菌的分离鉴定及其生长代谢特性
凌空1,翟磊1,2,姚粟1,宋振2,杨玉新2,程池1*
1(中国食品发酵工业研究院,北京,100015) 2(新疆中亚食品研发中心(有限公司),新疆 乌鲁木齐,830001)
从新疆传统发酵骆驼奶、马奶子和辣椒酱中分离筛选得到13株乳酸菌,通过16S rRNA基因序列和pheS基因序列系统发育学分析,结合表型特征确定乳酸菌的分类学地位。通过测定耐酸耐盐特性、产酸特性、降解亚硝酸盐能力、氨基酸脱羧酶活力及抑菌能力,筛选获得了3株具有潜在生产应用价值的乳酸菌,即LactobacilluspentosusCPC1 (CICC 6283)、LactobacilluspentosusCPC2 (CICC 6294)和LactobacilluskefiranofaciensCC3 (CICC 6287),为乳酸菌在果蔬发酵中的应用奠定了菌种基础。
新疆传统发酵食品;乳酸菌;分离鉴定;生长代谢特性
新疆传统发酵食品的制作和食用历史悠久,风味独特,营养丰富,而经过长时间的自然驯化,这些传统发酵食品中一些具有优良特性的乳酸菌保留了下来,在发酵食品过程中起到了重要作用[1]。乳酸菌因其对人体有独特的健康功效[2-4]引起了科学研究者们的广泛关注。对于乳酸菌的应用、益生功能和作用机制及其代谢组学、基因组学的研究是当前研究热点。在发酵蔬菜应用方面,乳酸菌被用来生产富含γ-氨基丁酸、B族维生素和乳酸钙的产品,而且其新功能不断被揭示[5-7]。
本研究采用分子生物学鉴定技术结合表型特征对新疆传统发酵骆驼奶、马奶子和辣椒酱中分离的乳酸菌进行鉴定,明确其分类学地位。在此基础上,从耐酸耐盐特性、产酸能力、抑菌能力、降解亚硝酸盐能力和氨基酸脱羧酶能力等方面对乳酸菌的生长代谢特征进行测定分析,筛选得到了3株具有潜在生产应用价值的乳酸菌,为乳酸菌在果蔬发酵中的应用奠定了基础。
1 材料与方法
1.1 材料与试剂
1.1.1 实验材料
新疆传统发酵食品包括新疆乌鲁木齐市乌鲁木齐县南山地区的传统发酵骆驼奶和马奶子,新疆巴音郭楞蒙古自治州博湖县发酵辣椒酱及新疆中亚食品研发中心发酵辣椒酱。
1.1.2 实验菌株
EscherichiacoliO157:H7 CICC 10907,Salmonellaentericasubsp.entericaserovarTyphiCICC 10871,StaphylococcusaureusCICC 10790和ListeriamonocytogenesCICC 21635。
1.1.3 实验试剂及设备
MRS培养基、胰蛋白胨大豆肉汤(Trypticase Soy Agar,TSA)、革兰氏染色试剂盒和生理生化鉴定管均购自北京陆桥技术有限公司;细菌基因组DNA提取试剂盒购于OMEGA公司;GoldView购自北京赛百盛基因技术有限公司;溶菌酶购自Sigma公司、蛋白酶购自Merk公司;TaqDNA聚合酶、dNTP、DL2000 marker购自天为时代生物有限公司;其他化学药品均为分析纯产品。
光学显微镜Olympus BH-2购于奥林巴斯有限公司;pH计FE20购于梅特勒-托利多仪器(上海)有限公司;紫外分光光度计7200购于尤尼科(上海)仪器有限公司;温度梯度PCR仪购于Biometra公司;恒温培养箱BHG-8082型购于上海一恒科学仪器有限公司。
1.2 实验方法
1.2.1 乳酸菌的分离纯化
利用梯度稀释涂布法对样品中乳酸菌进行分离,分别取25 mL的骆驼奶、马奶子和25 g的辣椒酱加入到225 mL无菌生理盐水中,用涡旋振荡器摇匀,制得10-1的稀释液,再从中吸取1 mL稀释液到9 mL无菌生理盐水中形成10-2的稀释液,依次制得10-3、10-4、10-5、10-6的稀释液。分别取100 μL稀释液涂布于添加了2% CaCO3的MRS培养基,置于37 ℃下培养48 h。挑取有溶钙圈的单菌落进行平板划线纯化[8-9],重复分离纯化3次,挑取单个菌落进行革兰氏染色,于100倍光学显微镜下观察菌体的形态学特征并记录。再挑取单菌落,在其表面滴加20%的过氧化氢溶液,观察有无气泡产生,并记录结果。分离纯化后菌株在斜面上连续传代2代后加入20%甘油进行保藏并编号。
1.2.2 乳酸菌分子生物学鉴定
通过分子生物学(16S rRNA基因序列测定及pheS基因序列分析)对分离纯化的菌株进行鉴定。
使用细菌基因组DNA提取试剂盒提取乳酸菌的基因组。利用引物27f (5’-AGAGTTTGATCCTGGCTCAG-3’)和1492r (5’-GGTTACCTTGTTACGACTT-3’)对16S rRNA基因进行扩增。PCR反应体系为:模板10×PCR buffer 5 μL、dNTP (2.5 mmol/L) 4 μL、模板 2 μL、TaqDNA 聚合酶1 μL、引物各1 μL,补充去离子水至50 μL。反应条件:94 ℃ 5 min,94 ℃ 50 s,52 ℃ 50 s,72 ℃ 50 s,33个循环,72 ℃ 7 min。PCR产物送交北京诺赛基因组研究中心有限公司进行测序。
利用引物21f (5’-CAYCCNGCHCGYGAYATGC-3’)和23r(5’-GGRTGRACCATVCCNGCHCC-3’)对pheS基因序列进行扩增。PCR反应体系同上,反应条件:95 ℃ 5 min,95 ℃ 1 min,46 ℃ 2 min 15 s,72 ℃ 1 min 15 s,3个循环;95 ℃ 1 min 15 s,46 ℃ 1 min 15 s, 72 ℃ 1 min 15 s,30个循环,72 ℃ 7 min。PCR产物送交北京诺赛基因组研究中心有限公司进行测序。
将测序结果在GenBank数据库中进行比对分析,以确定其与模式菌种的同源关系。确定并下载各模式菌种的有效序列后,采用ClustalX 1.83进行多序列比对后,使用MEGA 5.0软件中的邻接法(Neighbor-Joining)进行系统发育分析[10]。
1.2.3 乳酸菌耐酸试验
保藏的菌株经过甘油管转接液体培养基并传代2次后,按1%接种量分别接入pH2、3、4、5、6和pH7的MRS液体培养基中,37 ℃静置培养48 h,测定620 nm下吸光值(OD620),并记录结果[11]。
1.2.4 乳酸菌耐盐试验
保藏的菌株经过甘油管转接液体培养基并传代2次后,按1%接种量接入分别含2%、4%、6%、8%和10%(w/v)NaCl的MRS液体培养基中,37 ℃静置培养48 h,测定620 nm下吸光值(OD620),并记录结果[12]。
1.2.5 乳酸菌生长曲线及产酸能力测定
保藏的菌株经过甘油管转接液体培养基并传代2次后,按1%接种量接种于MRS液体培养基中培养,以不接种培养基作为对照,每隔2 h取样测定620 nm下吸光值(OD620)。同时每隔4 h取出部分发酵液通过pH计直接测定pH值[7, 13]。
1.2.6 乳酸菌亚硝酸盐降解能力测定
保藏的菌株经过甘油管转接液体培养基并传代2次后,按1%接种量接种于含125 μg/mL NaNO2的200 mL MRS液体培养基中37 ℃培养,每隔24 h定时取样测定NaNO2含量。参考GB/T 5009.33—2003中的盐酸萘乙二胺法进行测定,不接种的MRS培养基(含125 μg/mL NaNO2)作为空白对照[14]。
1.2.7 乳酸菌氨基酸脱羧酶试验
保藏的菌株经过甘油管转接液体培养基并传代2次后,接种于MRS培养基上,37 ℃培养24 h,挑取MRS培养基上生长的单菌落于3 mL无菌生理盐水中研磨,制备成0.5 McFarland悬液,分别滴入氨基酸脱羧酶试验的安培瓶中,每瓶3滴,并加无菌液体石蜡覆盖培养基表面,培养18~24 h后,观察试验管与对照管颜色变化,试验管为紫色,对照管为黄色,结果为阳性;试验管与对照管均为黄色,结果为阴性。
1.2.8 乳酸菌抑菌性研究
参考张佳等[15]的方法并作少许修改,采用滤纸片法测定目标菌株的抑菌性能。挑取新鲜培养的指示菌株(EscherichiacoliO157:H7 CICC 10907,Salmonellaentericasubsp.entericaserovarTyphi CICC 10871,StaphylococcusaureusCICC 10790和ListeriamonocytogenesCICC 21635),将菌悬液浓度调至0.5 McFarland并均匀涂布在TSA培养基上,然后用无菌镊子夹取无菌滤纸片放入已培养48 h的乳酸菌发酵上清液中,充分浸泡后,置于上述培养基中,37 ℃培养箱中培养48 h,记录结果。
2 结果分析
2.1 乳酸菌的分离纯化
从MRS平板上挑取13个有溶钙圈且外观不同的菌落,纯化后获得的菌株分别命名为CM1、CM2、CM3、CM4、CC1、CC2、CC3、CPC1、CPC2、CP1、CP2、CPZ1和CPZ2。革兰氏染色以及接触酶实验表明13株菌株均为革兰氏阳性,接触酶阴性菌株,符合乳酸菌的特征(表1)。
2.2 乳酸菌分子生物学鉴定
16S rRNA基因系统发育分析表明13株菌株均属于乳酸菌,其中菌株CC2和CC3与产马乳酒乳杆菌的同源性为99.86%,且与其他近种的同源性均在98.65%以下,则菌株CC2和CC3可鉴定为Lactobacilluskefiranofaciens。菌株CPZ2与消化乳杆菌的同源性为99.93%,且与其他近种的同源性均在98.65%以下,则菌株CPZ2可鉴定为Lactobacillusalimentarius,其余10株菌株只能鉴定到属水平(图1-A)。对于16S rRNA基因序列只能鉴定到属水平的菌株进行pheS基因序列系统发育分析。结果表明菌株CPZ1、CP1、CPC1和CPC2与戊糖乳杆菌的同源性为100%,可鉴定为Lactobacilluspentosus,CC1、CM1、CM2、CM3和CM4与瑞士乳杆菌的同源性为100%,可鉴定为Lactobacillushelveticus,而菌株CP2与耐乙酸片球菌的同源性为100%,鉴定为Pediococcusethanolidurans(图1-B)。分离鉴定的13株乳酸菌均已保藏于中国工业菌种保藏管理中心(CICC)(表1)。
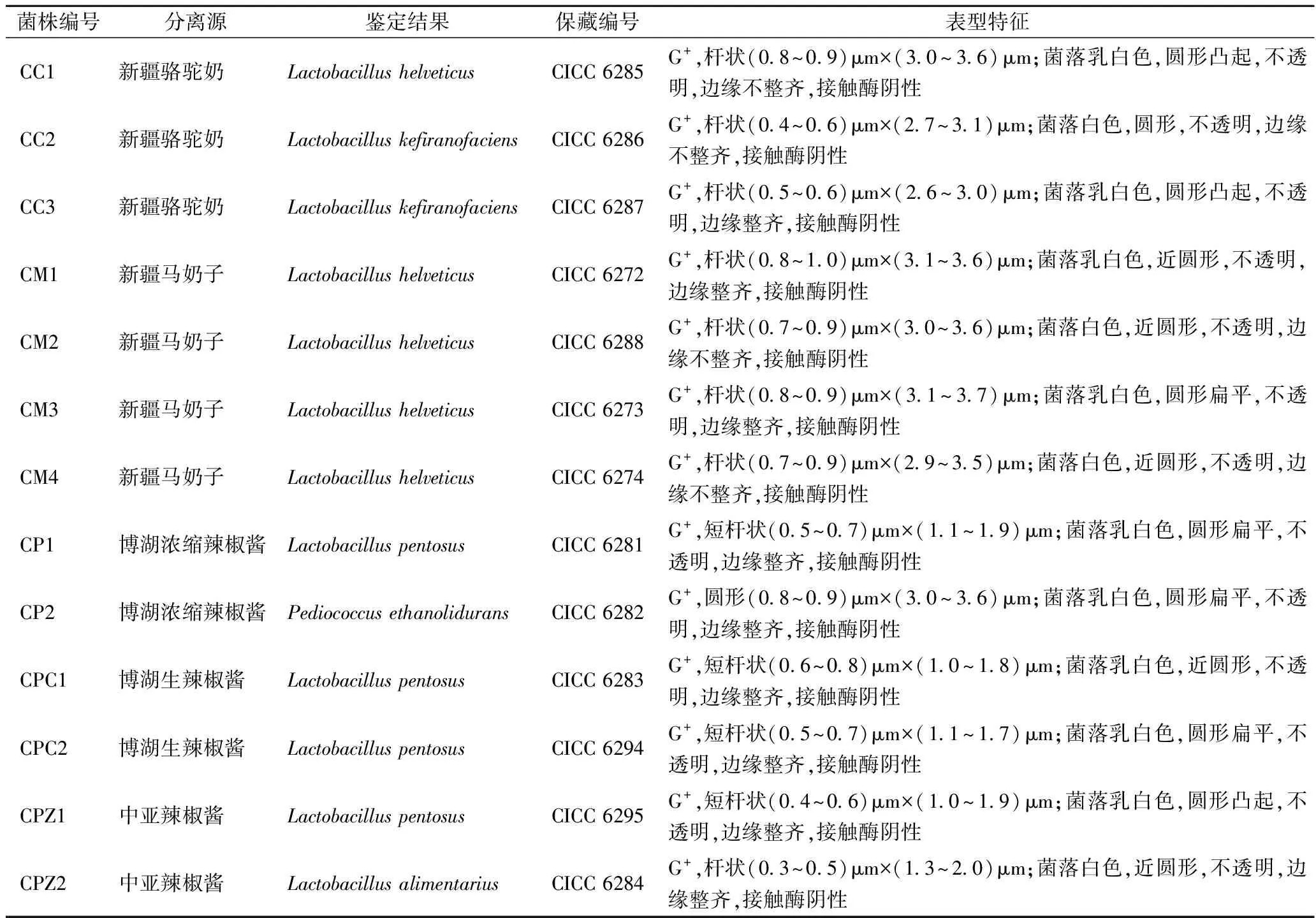
表1 分离鉴定的乳酸菌信息统计
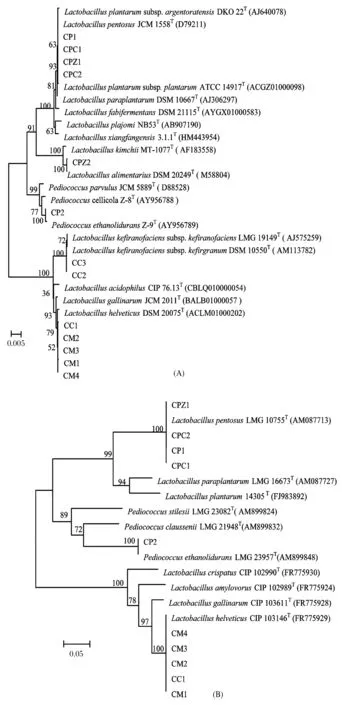
A-16S rRNA基因;B-pheS基因图1 分离鉴定的乳酸菌系统发育树Fig.1 The phylogenetic trees of isolated and identified lactic acid bacteria采用MEGA5.0软件,邻位连接法显示菌株与相关模式种系统发育树,进行1000次的相似度重复计算,图中发育树节点只显示Bootstrap值大于50%数值,上标的“T”表示模式菌株
2.3 乳酸菌耐酸耐盐实验
耐酸试验结果表明,随着pH值降低,乳酸菌的生长受到明显的抑制。其中菌株CC3、CPC1和CPC2展现了较强的耐酸能力,其中菌株CC1在pH 3条件下OD620值能达到0.16,而其他菌株只能达到0.05左右,其中菌株CPC2在pH 4的条件下的OD620值可以达到0.94。而菌株CP2耐酸能力最弱,不能在pH 4的环境下生长,在pH 5、6和pH7的环境中生长也较弱(图2-A)。
耐盐试验结果表明,乳酸菌的生长与NaCl浓度成反比,即随着NaCl浓度的升高,乳酸菌的生长受到明显的抑制(图2-B)。所有菌株在2%的NaCl下可以正常生长,但CM4、CP2和CPZ2的生长缓慢。而菌株CPC1的耐盐能力最强,在8%的NaCl条件下,OD620值可以达到0.55。菌株CC1、CC3、CPC1、CPC2和CM1能够耐受6%的NaCl。综合分析乳酸菌的耐酸和耐盐能力,选取菌株CC1、CC3、CPC1、CPC2和CM1进行后续生长代谢特性研究(图2-B)。

A-耐酸特性;B-耐盐特性图2 分离鉴定的乳酸菌耐酸耐盐特性 Fig.2 Acid and salt-resistance properties of isolated and identified lactic acid bacteria
2.4 乳酸菌的生长曲线和产酸能力测定

A-生长特性;B-产酸特性图3 分离鉴定的乳酸菌的生长和产酸特性 Fig.3 Growth and acid production of isolated and identified lactic acid bacteria
生长曲线测定表明,菌株CPC1、CC3和CPC2进入对数期较快,分别在8 h、10 h和10 h最早进入对数期,分别在16 h、32 h和34 h进入稳定期,而CM1在16 h最晚进入对数期(图3-A)。产酸能力测定结果表明,菌株CPC1产酸速度最快,产酸能力最大,pH值可以降到3.62,菌株CC3和CPC2产酸速度次之,pH值可以降到3.83(图3-B)。由此可见,菌株进入对数期越快,其产酸速率也较快。
2.5 乳酸菌亚硝酸盐降解能力测定
随着培养时间的延长,菌株降解的亚硝酸盐越多。培养24 h后,CC3、CPC1和CPC2的降解率分别可达到92.61%、94.82%和98.84%,而CC1和CM1的降解率只有34.35%和24.26%。培养48 h后,除CC1和CM1的降解率达到82.73%和81.84%外,其余3株降解率均可以达到100%(表2)。

表2 分离鉴定的乳酸菌表型特性
注:“a”表示菌株培养24 h后的亚硝酸盐降解率,“b”表示菌株培养48 h后的亚硝酸盐降解率;“+”表示阳性;“-”表示阴性。
2.6 乳酸菌氨基酸脱羧酶试验
经过氨基酸脱羧酶试验,菌株CM1对精氨酸双水解和鸟氨酸脱羧酶活性显阳性,剩余菌株对4种氨基酸脱羧酶活性均显阴性(表2)。
2.7 乳酸菌抑菌能力测定
由抑菌能力实验结果可知,5株乳酸菌均能够抑制菌肠炎沙门氏菌和大肠杆菌,其中CPC1和CPC2的抑菌作用最大,对于肠炎沙门氏菌的抑菌直径可达到24.2 mm和26.0 mm,对于大肠杆菌的抑菌直径可达到16.3 mm和13.3 mm。仅CC1不能抑制金黄色葡萄球菌,其中CPC2抑制作用最大,抑菌直径为19.4 mm,菌株对单增李斯特菌的抑菌作用较弱,而CC3对4种病原菌都有抑制作用(表2)。
3 结论
通过分子生物学鉴定,结合表型特征确定乳酸菌的分类学地位。从新疆传统发酵食品中共筛选到2个属,13株乳酸菌。其中瑞士乳杆菌5株,戊糖乳杆菌4株,产马乳酒乳杆菌2株,消化乳杆菌和耐乙酸片球菌各1株,并将这些乳酸菌资源保藏于中国工业微生物菌种保藏管理中心(CICC),丰富了新疆传统发酵食品的乳酸菌资源。从菌株来源看,新疆马奶子中主要含有瑞士乳杆菌,而骆驼奶中既含有瑞士乳杆菌又含有产马乳酒乳杆菌。其中瑞士乳杆菌也是由熊素玉等[10]从新疆酸马奶中分离的主要乳酸菌,而产马乳酒乳杆菌的研究报道较少,是今后研究的重点。辣椒酱中都含有戊糖乳杆菌,而博湖浓缩辣椒酱中还含有耐乙酸片球菌,来自中亚食品的辣椒酱除了戊糖乳杆菌外,还含有消化乳杆菌。由此可见,不同来源的辣椒酱中既含有相同乳酸菌又各具特色(图4)。因此本研究对于新疆传统发酵食品中乳酸菌资源的收集积累具有重要意义。

图4 分离鉴定的乳酸菌的种类和来源Fig.4 Classification and origin of isolated and identified lactic acid bacteria
乳酸菌对低酸环境和渗透压的耐受性也是作为发酵剂和益生菌的必要条件[2]。纪晓燚[7]等以乳酸菌在不同pH条件下的适应性为指标,试验筛选出耐酸较好的菌株在pH 4条件下的OD620在0.5左右,并将其应用于发酵。BEGANOVIC[11]对筛选到的菌株进行4%~6% NaCl条件下的耐盐试验,认为优良菌株的OD600值在0.4~0.6之间。本研究中筛选得到的菌株CC3、CPC1和CPC2在pH 4条件下的OD620均在0.5以上,菌株CPC1在盐浓度为6%条件下的OD620为1.09,展现了良好的耐酸和耐盐能力,HWANHLEM[16]等认为乳酸可以导致革兰氏阴性病原菌外膜的损伤,引起病原菌LPS层的破坏从而导致的亚致死损伤,试验选取降低pH能力作为筛选指标,得到产酸可以使pH值降至pH4.3~4.6的菌株应用于发酵中。AMMOR[17]等认为乳酸菌快速产酸形成的低酸环境可以抑制有害菌群的繁殖从而提升产品的安全性及货架期。而本实验中筛选得到的菌株产酸后均可以使pH值降至4.30以下,为后续的果蔬发酵试验奠定了基础。
根据GB/T 2762—2012食品中污染物限量规定,蔬菜及其制品腌渍蔬菜的亚硝酸盐(以NaNO2计)含量不大于20 mg/kg,摄取过量的亚硝酸盐会引起致癌、抵抗甲状腺素和引起智障等危害[18]。FRIEDRICH-KARL[19]等在关于德国常见传统食品的安全挑战研究中就提到生物胺的危害性,生物胺是菌株利用氨基酸在氨基酸脱羧酶的作用下产生的,如果健康人群摄入大量生物胺,或易感人群摄入少量生物胺,生物胺能进入机体各组织系统,导致肾上腺素和胃酸过量分泌、心跳加快、血糖含量增加或血压升高等症状[20]。YOSEP[21]等在分离于韩国泡菜中乳酸菌的安全性评价中把是否产生生物胺作为考察指标之一。所以果蔬发酵中亚硝酸盐及生物胺的含量也是考虑的重要因素。因此筛选降解亚硝酸能力强和不产生物胺的菌株作为发酵剂能够降低最终产品中亚硝酸盐和生物胺的含量,提高产品的安全性。菌株CC3、CPC1和CPC培养48 h后亚硝酸盐的降解率可达100%,高于目前报道的乳酸菌[22]。而且菌株CC3、CPC1和CPC2的4种氨基酸脱羧酶均为阴性,不会产生生物胺。同时对4种有害菌种均有抑菌作用,将其应用在果蔬发酵中,能够进一步提高产品的安全性。
综上所述,本研究筛选得到了13株来自新疆传统发酵食品的乳酸菌,对于新疆乳酸菌资源的收集和积累具有重要意义。通过功能筛选,获得了3株具有潜在生产价值的乳酸菌,即LactobacilluspentosusCPC1 (CICC 6283)、LactobacilluspentosusCPC2 (CICC 6294)和LactobacilluskefiranofaciensCC3 (CICC 6287),为乳酸菌在果蔬发酵中的应用奠定了菌种基础。
[1] 拉提帕·艾尔肯,唐雪,新华·那比.新疆传统发酵驼乳中分离出的一株乳酸菌的分子生物学鉴定[J].新疆医科大学学报, 2014,37(2): 155-159.
[2] ARGYRI A A,ZOUMPOPOULOU G,KARATZAS K A, et al.Selection of potential probiotic lactic acid bacteria from fermented olives byinvitrotests [J].Food microbiology,2013,33(2):282-291.
[3] BAO Y,ZHANG Y,ZHANG Y, et al.Screening of potential probiotic properties ofLactobacillusfermentumisolated from traditional food products [J].Food Control,2010, 21(5): 695-701.
[4] 高娃.四川部分地区泡菜中乳酸菌的分离鉴定[D].呼和浩特:内蒙古农业大学,2010.
[5] 赖婷,刘汉伟,张名位,乳酸菌发酵对果蔬中主要活性物质及其生理功能的影响研究进展[J].中国酿造,2015,34(3): 1-4.
[6] 谢明勇,熊涛,关倩倩.益生菌发酵果蔬关键技术研究进展[J].中国食品学报,2014,14(10):1-9.
[7] 纪晓燚.优良乳酸菌的筛选及其发酵蔬菜的应用研究[D].杭州:浙江大学, 2014.
[8] 杨洁,张文亮,邹建军,等.新疆传统酸奶中乳酸菌的筛选鉴定及菌相分析[J].食品工业科技,2015,36(1):324-327.
[9] XIONG T, LI X, GUAN Q,et al. Starter culture fermentation of Chinese sauerkraut: Growth, acidification and metabolic analyses [J]. Food Control, 2014, 41(2):122-127.
[10] 熊素玉.酸马奶中乳酸菌的分离鉴定及其生物学特性的研究[D].乌鲁木齐:新疆农业大学,2007.
[11] 胡书芳,王雁萍,洪爱俊,等.自然发酵酸菜中乳酸菌的分离鉴定及其生理特性研究[J].安徽农业科学,2009, 37(15): 6 896-6 898.
[12] BEGANOVIC J,KOS B,LEBOS PAVUNC A,et al.Traditionally produced sauerkraut as source of autochthonous functional starter cultures [J]. Microbiological research, 2014, 169(7-8): 623-632.
[13] 杨俊俊.西藏牦牛奶渣中微生物的分离鉴定及优良乳酸菌的筛选[D].杨凌:西北农林科技大学,2014.
[14] 高书锋,陈丁,贺莫许等.亚硝酸盐降解菌的分离鉴定及其降解特性[J]. 环境科学与技术, 2011(S2): 37-41.
[15] 张佳,王莹,张峰等.滤纸片法测定黄花蒿提取物对霉菌的抑制活性[J]. 湖北农业科学,2009,48(5):1 153-1 154.
[16] HWANHLEM N,BURADALENG S,WATTANACHANT S,et al.Isolation and screening of lactic acid bacteria from Thai traditional fermented fish (Plasom) and production of Plasom from selected strains [J].Food Control,2011, 22(3-4): 401-407.
[17] AMMOR M S, MAYO B. Selection criteria for lactic acid bacteria to be used as functional starter cultures in dry sausage production: An update [J]. Meat science, 2007, 76(1): 138-146.
[18] 管世敏.降解亚硝酸盐乳酸菌的分离筛选及其在泡菜发酵中的应用研究[D].上海:上海师范大学,2009.
[19] FRIEDRICH-KARL LÜCKE, PETER Z. Food safety challenges associated with traditional foods in German-speaking regions[J]. Food Control, 2014, 43(5):217-230.
[20] 姜维.一株耐盐性高效生物胺降解新菌的筛选分类鉴定及应用研究[D].青岛:中国海洋大学,2014.
[21] YOSEP J, HANNAH K, HYUNJOON P,et al。Functionality and safety of lactic bacterial strains from Korean kimchi[J]. Food Control, 2013,31(3): 467-473.
[22] 吴慧昊,牛锋,陈珊珊等.高效降亚硝酸盐乳酸菌的驯化复筛及菌株鉴定[J]. 食品科学,2016,37(19):160-165.
Isolation, identification and characterization of lactic acid bacteria from traditionally fermented foods in Xinjiang
LING Kong1,ZHAI Lei1,2,YAO Su1,SONG Zhen2,YANG Yu-xin2,CHENG Chi1*
1(China National Research Institute of Food and Fermentation Industries, Beijing 100015, China) 2(Xinjiang Central Asia Food Research and Development Centre, Urumchi 830001, China)
Thirteen strains of lactic acid bacteria were isolated from traditionally fermented foods including camel milk, mare milk and chilli sauce in Xinjiang. These lactic acid bacteria were identified by phylogenetic analyses of 16S rRNA gene and pheS gene, combined with phenotypic characteristics. Based on the abilities of acid- and salt-resistance, acid production, nitrite degradation, activity of amino acids decarboxylase and antimicrobial properties, the lactic acid bacteria with potential for application were chosen. As a result, three lactic acid bacteria designated asLactobacilluspentosusCPC1 (CICC 6283),LactobacilluspentosusCPC2 (CICC 6294) andLactobacilluskefiranofaciensCC3 (CICC 6287) were obtained and laid a foundation for vegetable fermentation application.
naturally fermented food in Xinjiang; lactic acid bacteria (LAB); isolation and identification; growth metabolism
10.13995/j.cnki.11-1802/ts.201703022
硕士研究生(程池教授为通讯作者,E-mail:cheng100027@163.com)。
国家微生物资源平台专项(No. NIMR2016-4);自治区科技支疆项目(2016E02021);中国食品发酵工业研究院科技发展基金(博士基金)项目(No. 2015KJFZ-BS-04)
2016-09-12,改回日期:2016-11-29
