乳腺癌MCF-7细胞p21WAF1/CIP1启动子区HDAC1高功能结合位点的研究
2017-03-17周伟强
邹 丹, 周伟强
(沈阳医学院1.病理生理学教研室、2.辽宁省环境污染与微生态重点实验室,辽宁 沈阳 110034)
◇论 著◇
乳腺癌MCF-7细胞p21WAF1/CIP1启动子区HDAC1高功能结合位点的研究
邹 丹1, 周伟强2
(沈阳医学院1.病理生理学教研室、2.辽宁省环境污染与微生态重点实验室,辽宁 沈阳 110034)
目的 研究乳腺癌MCF-7细胞中组蛋白去乙酰化酶1(histone deacetylases 1,HDAC1)募集于p21WAF1/CIP1启动子区调控其转录活性的特异性结合位点。方法 将处于对数生长期的乳腺癌MCF-7细胞在无血清培养基中饥饿24 h后,分别用20 μmol·L-10.88 μL SAHA(S 组)、0.625 nmol·L-110 μL Leptin(L组)处理24 h,对照组(B组)细胞培养在完全型RPMI 1640培养基中。各组细胞裂解液与HDAC1抗体孵育,收集纯化结合HDAC1抗体的DNA片段,应用Real-time PCR法检测p21WAF1/CIP1启动子区从TSS到其上游(+2~-4 000 bp) f1~f10片段的DNA相对表达量并用2-ΔΔCT法分析。结果 B组中,HDAC1抗体在p21WAF1/CIP1启动子区f1、f8片段有高亲和力,f8片段达最高。S组中,HDAC1抗体与p21WAF1/CIP1启动子区f1~f10片段结合量明显低于对照组,f8片段达最低,而在L组此片段与HDAC1抗体结合量达最大值。结论 乳腺癌MCF-7细胞增殖过程中, HDAC1可被招募至p21WAF1/CIP1启动子区, 该启动子区上游-2 800 bp至-3 200 bp DNA片段是与HDAC1高度结合的靶功能区。
乳腺癌; MCF-7细胞;p21WAF1/CIP1;组蛋白去乙酰化酶1;辛二酰苯胺异羟肟酸;瘦素
乳腺癌的发生发展与多种肿瘤细胞增殖、凋亡、侵袭、转移的蛋白有关。组蛋白去乙酰化酶1(histone deacetylases 1,HDAC1)使组蛋白去乙酰化,增加组蛋白与DNA双链的亲和性,可与一些调控肿瘤发生发展的基因启动子区结合而抑制这些基因的转录。HDAC抑制剂(histone deacetylase inhibitor, HDACi)是一类可以抑制组蛋白去乙酰化酶活性、维持组蛋白乙酰化水平的化合物,可促使肿瘤细胞生长停滞并诱发细胞凋亡。辛二酰苯胺异羟肟酸 (suberoylanilide hydroxamic acid,SAHA)是目前已知最经典的HDACi之一, 主要用于皮肤T细胞淋巴瘤的治疗。SAHA可以通过上调p21WAF1/CIP1引起细胞周期阻滞于G1期而抑制细胞增殖[1]。
p21WAF1/CIP1是细胞周期依赖性蛋白激酶抑制剂,可使细胞周期停滞、促进细胞凋亡并抑制癌细胞的转移和侵袭。乳腺癌p21WAF1/CIP1的表达与细胞分化程度无关,与淋巴结转移显著负相关,p21WAF1/CIP1的高表达有抑制肿瘤转移的作用[2]。
瘦素(Leptin)是肥胖基因的表达产物,主要由脂肪细胞合成和分泌,包含167个氨基酸,通过与瘦素受体OB-Rb结合发挥作用。Leptin血清水平与脂肪储存量成正比[3],研究表明:随着女性体内Leptin水平增加,其患高恶性、低分化且预后不良的乳腺癌几率大大增加[4]。本研究的前期工作证实Leptin处理后的小鼠乳腺癌4T1细胞p21WAF1/CIP1 mRNA及蛋白表达水平远远低于Leptin未处理组细胞[5]。
本实验旨在找出HDAC1募集于p21WAF1/CIP1启动子区调控其转录活性的具体作用位点,明确SAHA及Leptin在调节p21WAF1/CIP1启动子功能过程中的分子机制。
1 材料与方法
1.1 材料 人乳腺癌细胞株MCF-7(美国ATCC细胞库),SAHA、Leptin(Sigma), 完全型RPMI 1640培养液、改良型RPMI 1640培养液、磷酸盐缓冲液1X(Thermo), 0.25%胰酶-EDTA(Gibco), PierceTMAgarose ChIP试剂盒(Thermo), HDAC1抗体(Abcam), Power SYBR®Green PCR Master Mix(Life technologies),p21WAF1/CIP1f1~f10引物(上海生工生物工程有限公司), 超净工作台、CO2恒温细胞培养箱(Thermo), Biofuge 28RS低温高速离心机(Heraeus), 倒置显微镜(Olympus), SK-D1807-E 摇床(Scilogex), 恒温混匀器(Eppendorf), 7500 Real-Time PCR仪(Life Technologies)。
1.2 方法
1.2.1 细胞培养 将MCF-7细胞接种在完全型RPMI 1640培养液(10%胎牛血清,青霉素、链霉素100 kU·L-1)中,37℃、5% CO2孵箱饱和湿度条件下贴壁传代培养,每2~3天换液并传代1次。0.25%胰酶-EDTA消化。经2~3次传代,取对数生长期的细胞等量分成3组,分别接种于完全型RPMI 1640培养液中,24 h后,再将细胞置于改良型RPMI 1640培养液中进行同步化处理24 h,此后将3组细胞分别定义为SAHA组(S组,即完全型RPMI 1640培养液10 mL+20 μmol·L-1SAHA 0.88 μL)、Leptin组(L组,即完全型RPMI 1640培养液10 mL+0.625 nmol·L-1Leptin 10 μL),Basal对照组(B组,即加等量完全型RPMI 1640培养液)。
1.2.2 应用染色质免疫共沉淀技术(chromatin-immunoprecipitation,ChIP)处理样品 MCF-7细胞的甲醛交联及细胞团的分离;细胞溶解及MNase 消化,每组样品各得到50 μL的MNase消化产物,留取5 μL -20℃保存作为内对照,45 μL进行下一步的免疫沉淀处理,免疫沉淀过程中需向每个ChIP样品加入HDAC1 抗体10 μg,实验步骤均按操作手册完成。最终3组MCF-7细胞均获得ChIP得到的DNA及各自的内对照。
1.2.3 Real-time PCR检测各组p21WAF1/CIP1f1~f10片段的DNA相对表达 PCR反应体系为25 μL:DNA 1.5 μL,上下游引物各0.75 μL,PCR Master Mix 12.5 μL, ddH2O 9.5 μL。反应条件为:50℃ 2 min,95℃ 10 min预变性, 95℃ 15 s, 60℃ 1 min, 共40个循环。每个样品3个复孔,采用2-ΔΔCT法计算各组DNA 的相对表达量。
p21WAF1/CIP1启动子区的10个片段的引物序列为: f1:Forward 5′-TCC TCC TGG AGA GTG CCA AC-3′,Reverse 5′-TTG GTG CGC TGG ACA CAT TT-3′;f2:Forward 5′-TTC CCG GAA GCA TGT GAC AA-3′,Reverse 5′-GCA CCT GGA GCA CCT AGA CAC C-3′;f3:Forward 5′-CCC GTT TCC CCA GCA GTG TA-3′,Reverse 5′-GCC AGG AAG GGG AGG ATT TG-3′;f4:Forward 5′-AGG CCA AGG GGG TCT GCT AC-3′,Reverse 5′-CGG GGA GGA CAG GCT TCT TT-3′;f5:Forward 5′-TGA AAG CAG AGG GGC TTC AA-3′,Reverse 5′-ACC ATC CAA AGG GCT GGT TG-3′;f6:Forward 5′-TGT CCT TGG GCT GCC TGT TT-3′,Reverse 5′-AGC CCT GTC GCA AGG ATC TG-3′;f7:Forward 5′-TTC TGC AGC CAC CAC TGA GC-3′,Reverse 5′-GTG GAG CAG CAT GGG GTA GG-3′;f8:Forward 5′-CCC ACC TCA GCC ACC TGA AT-3′,Reverse 5′-GGG CAG ATC ACA GGG TCA GG-3′;f9:Forward 5′-AGT GGG CAC ATT TAG ACA TAG CAG GT-3′,Reverse 5′-CCT CCC GGT CAT GCC TTT C-3′;f10:Forward 5′-GTC AGG TGC CAC TGG GGT CT-3′,Reverse 5′-CGG TCC CCT GTT TCA ATG CT-3′。p21WAF1/CIP1启动子区+2~-4 000 bp 10个选定区域f1~f10(Fig 1)。
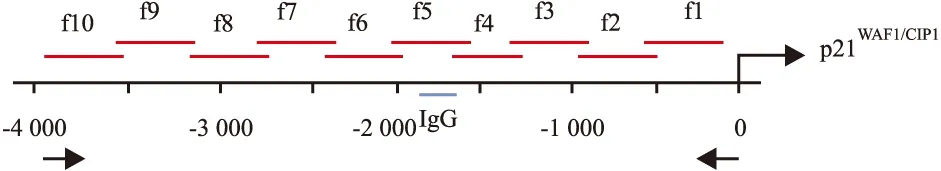
Fig 1 Schematic diagram of HDAC1 response element forregulating transcriptional function of p21WAF1/CIP promoter
2 结果
2.1 B组MCF-7细胞p21WAF1/CIP1启动子区f1~f10片段HDAC1高功能结合位点筛选 B组MCF-7细胞经HDAC1抗体Chip处理后,45 μL样品与5 μL内对照的DNA分别与f1~f10引物作用进行Real-time PCR扩增,所得CT值之差为该样品在p21启动子区各自片段的ΔCT值,再以f5片段为对照,其它片段的ΔCT值与其相减,得到B组样品在p21启动子区f1~f10片段各自ΔΔCT值,采用2-ΔΔCT法计算各片段DNA的相对表达量。B组f1、f2、f6、f7、f8片段DNA的相对表达量(1.88±0.18、1.68±0.07、1.35±0.13、1.49±0.09、2.11±0.26)较f5片段相对表达量(1±0)明显升高(P<0.01),f8片段达最高值(Fig 2)。
2.2 SAHA作用下MCF-7细胞p21WAF1/CIP1启动子区f1~f10片段HDAC1高功能结合位点筛选 S组MCF-7细胞DNA组分经HDAC1抗体ChIP处理后,进行Real-time PCR检测。结果表明:S组MCF-7细胞的p21WAF1/CIP启动子f1~f10片段 DNA的相对表达量(0.36±0.03、0.23±0.04、0.39±0.13、0.44±0.03、0.27±0.03、0.27±0.02、0.40±0.07、0.19±0.02、0.34±0.02、0.45±0.07)较B组对应片段相对表达量(1±0)均明显降低(P<0.01),f8片段达最低值(Fig 3)。
2.3 Leptin作用下MCF-7细胞p21WAF1/CIP1启动子区f1~f10片段HDAC1高功能结合位点筛选
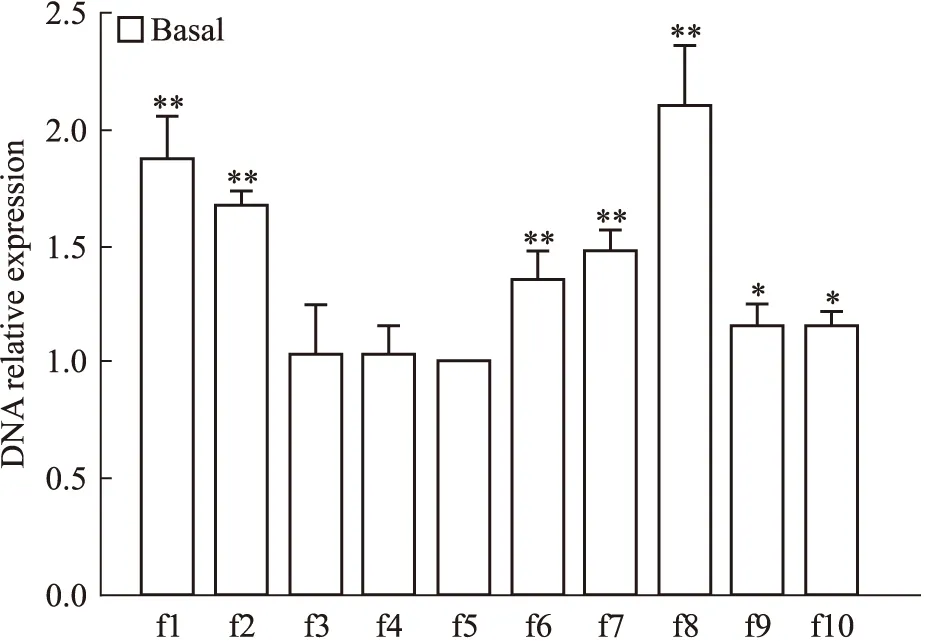
Fig 2 Screening of high function bind site of HDAC1 among f1~f10 fragments of p21WAF1/CIP promotor region in basal group MCF-7 cells
*P<0.05,**P<0.01vsf5 fragment
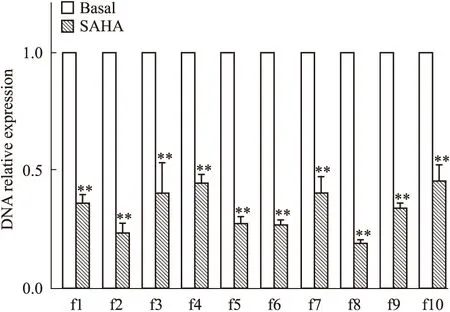
Fig 3 Screening of high function bind site of HDAC1 among f1~f10 fragments of p21WAF1/CIP promotor region in MCF-7 cells in SAHA group
**P<0.01vsbasal group
将Leptin处理后的MCF-7细胞的DNA组分经ChIP实验后进行Real-time PCR检测。结果表明:L组细胞p21WAF1/CIP启动子区f1~f10片段结合HDAC1抗体的DNA相对表达量(5.54±0.17、2.01±0.39、3.59±1.15、5.43±0.48、4.49±0.16、4.14±0.49、4.79±0.66、6.68±0.57、5.02±0.31、5.94±0.45)较B组对应片段相对表达量(1±0)均明显升高(P<0.01),f8片段达最高值(Fig 4)。
3 讨论
乳腺癌是女性恶性肿瘤之首,其中0.7~0.8的患者为雌激素受体(estrogen receptor,ER)阳性[6],因此选择ER阳性的MCF-7乳腺癌细胞系探索乳腺癌的发病机制及治疗药物尤为重要。
表观遗传改变是基因DNA序列不发生变化,但基因表达却发生了可遗传的改变。组蛋白乙酰化及去乙酰化等共价修饰是表观遗传学的主要研究内容,现已逐渐成为乳腺癌研究和治疗的热点[7]。在乳腺癌细胞增殖过程中,细胞增殖信号在p21WAF1/CIP1的启动子区招募HDAC1转录因子,通过组蛋白去乙酰化表观遗传修饰影响p21WAF1/CIP表达。SAHA通过抑制此过程而上调p21WAF1/CIP1基因的转录。Leptin通过激活HDAC1的活性使核心组蛋白去乙酰化,从而抑制p21WAF1/CIP1的表达,最终促进细胞周期从G1期到S期的进程[8]。
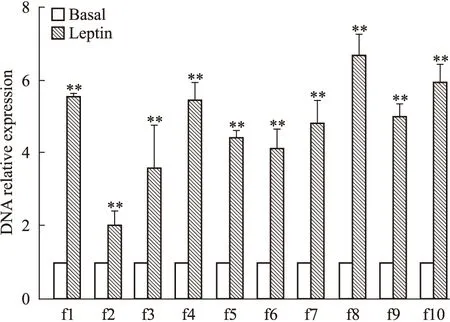
Fig 4 Screening of high function bind site of HDAC1 among f1~f10 fragments of p21WAF1/CIPpromotor region in MCF-7 cells in Leptin group
**P<0.01vsbasal group
ChIP可真实、完整地反映结合在DNA序列上的靶蛋白的调控信息, 是目前基于全基因组水平研究DNA-蛋白质相互作用的标准实验技术。本实验利用这一方法研究发现: B组 MCF-7细胞p21WAF1/CIP基因转录起始点近端启动子调节区f1和远端调节区f8片段结合HDAC1抗体的DNA相对表达量较其它区域明显升高。这表明在细胞增殖过程中,HDAC1可被招募至p21WAF1/CIP启动子区,促使组蛋白去乙酰化,增强组蛋白与DNA 的亲和力,抑制p21WAF1/CIP基因转录。
SAHA作用下的MCF-7细胞,与p21WAF1/CIP启动子区DNA结合的HDAC1抗体明显减少,这表明作用于p21WAF1/CIP启动子区的HDAC1明显减少,且在f8片段(-2 800 bp至-3 200 bp)处受SAHA影响HDAC1与p21WAF1/CIP启动子区DNA的结合量最少。SAHA作为一种广谱的HDACi,可抑制HDAC1的活性,从而提高组蛋白乙酰化的程度。高乙酰化的组蛋白可提高相关基因(如p21WAF1/CIP)的转录水平,最终表现为肿瘤细胞的增殖抑制、分化和凋亡[9]。另有研究报道:SAHA可降低HDAC1的表达而抑制癌组织的生长[10-11]。
Leptin可通过调控细胞周期、凋亡、影响细胞外环境等多重机制促进雌激素受体阳性的MCF-7乳腺癌细胞增殖与转移[12-14]。本实验中:Leptin作用下的MCF-7细胞,与p21WAF1/CIP启动子区DNA结合的HDAC1抗体明显增多,表明作用于p21WAF1/CIP启动子区的HDAC1明显增多,且在f8片段处受Leptin影响HDAC1与p21WAF1/CIP启动子区DNA的结合量最多。这一实验现象表明:在乳腺癌发生、发展过程中,某些细胞增殖信号招募HDAC1于p21WAF1/CIP启动子特定区域,调控Leptin对p21WAF1/CIP基因转录的作用。
综上所述,本研究通过ChIP技术筛选出乳腺癌MCF-7细胞p21WAF1/CIP1启动子区的HDAC1高功能结合位点,进而通过对SAHA及Leptin处理过的乳腺癌MCF-7细胞进行ChIP检测,明确p21WAF1/CIP启动子区-2 800 bp至-3 200 bp区域可能是与HDAC1高度结合的功能活跃区。此研究发现丰富了SAHA和Leptin在调节p21WAF1/CIP1启动子功能过程中的分子机制,更为抗乳腺癌的药物研发及靶向治疗奠定了理论和实验基础。
(致谢:衷心感谢沈阳医学院辽宁省环境污染与微生态重点实验室为本实验顺利完成提供的实验设备,感谢实验室全体工作人员对本实验的大力协助。)
[1] Lee Y J, Won A J, Lee J, et al. Molecular mechanism of SAHA on regulation of autophagic cell death in tamoxifen-resistant MCF-7 breast cancer cells[J].IntJMedSci,2012,9(10):881-93.
[2] Thor A D, Liu S, Moore D H, et al.p21(WAF1/CIP1) expression in breast cancers: associations with p53 and outcome[J].BreastCancerResTreat, 2000, 61(1):33-43.
[3] Grossmann M E,Ray A, Nkhata K J, et al. Obesity and breast cancer: status of leptin and adiponectin in pathological processes[J].CancerMetastasisRev, 2010, 29(4): 641-53.
[4] Hancke K,Grubeck D,Hauser N,et al. Adipocyte fatty acid-binding protein as a novel prognostic factor in obese breast cancer patients[J].BreastCancerResTreat, 2010, 119(2): 367-77.
[5] Zhou W, Guo S, Gonzalez-Perez R R. Leptin pro-angiogenic signature in breast cancer is linked to IL-1 signalling[J].BrJCancer, 2011, 104(1):128-37.
[6] Chargari C,Toillon R A,Macdermed D, et al. Concurrent hormone and radiation therapy in patients with breast cancer: what is the rationale[J]?LancetOncol,2009,10(1):53-60.
[7] Connolly D, Yang Z, Castaldi M,et al. Septin 9 isoform expression, localization and epigenetic changes during human and mouse breast cancer progression[J].BreastCancerRes, 2011,13(4):R76.
[8] Gonzalez-Perez R R,Xu Y,Guo S,et al. Leptin upregulates VEGF in breast cancer via canonic and non-canonical signalling pathways and NF-kappaB/HIF-1alpha activation[J].CellSignal, 2010, 22(9):1350-62.
[9] Fiskus W,Hembruff S L,Rao R,et al. Co-treatment with vorinostat synergistically enhances activity of Aurora kinase inhibitor against human breast cancer cells[J].BreastCancerResTreat,2012,135(2): 433-44.
[10]Jin J S,Tsao T Y,Sun P C,et al.SAHA inhibits the growth of colon tumors by decreasing histone deacetylase and expression of cyclin D1 and survivin[J].PatholOncolRes,2012,18(3):713-20.
[11]韩 翰,王 敏.SAHA和TRAIL联合使用对乳腺癌雌激素受体阳性细胞MCF-7生长的影响[J].中国药理学通报,2016,32(2):223-8.
[11]Han H, Wang M. Effects of combination treatment with SAHA and TRAIL on ER positive breast cancer cell MCF-7[J].ChinPharmacolBull, 2016,32(2): 223-8.
[12]Perera C N,Chin H G,Duru N,et al. Leptin-regulated gene expression in MCF-7 breast cancer cells:mechanistic insights into leptin-regulated mammary tumor growth and progression[J].JEndocrinol,2008,199(2):221-33.
[13]Strong A L,Ohlstein J F,Biagas B A, et al.Leptin produced by obese adipose stromal/stem cells enhances proliferation and metastasis of estrogen receptor positive breast cancers[J].BreastCancerRes,2015,17:112.
[14]冯秀艳,韩 翰,周伟强.SAHA 在Leptin 诱导的乳腺癌MCF-7细胞增殖过程中的调控作用[J].中国药理学通报,2016,32(4):503-8.
[14]Feng X Y, Han H, Zhou W Q. Regulation of SAHA on cell proliferation induced by leptin in breast cancer cell line MCF-7[J].ChinPharmacolBull, 2016, 32(4): 503-8.
A study on high function binding site of HDAC1 in p21WAF1/CIP1promoter region in breast cancer MCF-7 cells
ZOU Dan1, ZHOU Wei-qiang2
(1.DeptofPathophysiology; 2.KeyLaboratoryofEnvironmentalPollutionandMicroecologyofLiaoningProvince,ShenyangMedicalCollege,Shenyang110034,China)
Aim To investigate the specific binding sites that HDAC1 can be recruited to regulate the transcriptional activity of p21WAF1/CIP1promoter in the breast cancer MCF-7 cells.Methods The breast cancer MCF-7 cells in logarithmic growth phase were starved with FBS free medium for 24 hours, and treated with 20 μmol·L-1SAHA(S group) or 0.625 nmol·L-1Leptin(L group) for 24 hours, and the cells that were cultured in the complete RPMI 1640 medium without any treatment were assigned as control group(B group). The DNA-ChIP was followed the manufacturer′s protocol for the assay. The cell lysis was prepared and incubated with anti-HDAC1 antibody overnight at 4℃. DNA fragments binding anti-HDAC1 antibody were gathered and purified. The relative expression level of DNA fragments from TSS to the upstream of the p21WAF1/CIP1promoter region(+2~-4 000 bp) binding with antibody was detected by Real-time PCR and analyzed by 2-ΔΔCTmethod.Results In B group, HDAC1 had high affinity with the f1 and f 8 fragments of p21WAF1/CIP1promoter compared to the other fragemts, and showed the highest affinity with the f8 fragment. In S group, the binding ability of HDAC1 to the f1~f10 fragment of p21WAF1/CIP1promoter was significantly lower than that of the control. The binding activity of HDAC1 to f8 fragment was the lowest, while reversing to reach the peak after leptin treatment.Conclusions HDAC1 can be recruited to p21WAF1/CIP1promoter by the cell proliferation signal during the proliferation of breast cancer MCF-7 cells. The DNA fragment from -2 800 to -3 200 bp in the upstream of p21WAF1/CIP1promoter is the target functional region for the binding with HDAC1.
breast cancer;MCF-7 cell;p21WAF1/CIP1;HDAC1;suberoylanilide hydroxamic acid;leptin
时间:2017-3-4 11:49
http://kns.cnki.net/kcms/detail/34.1086.R.20170304.1149.012.html
2016-09-18,
2016-12-05
国家自然科学基金资助项目(No 81172509);辽宁省自然科学基金资助项目(No 201602735);沈阳市科技计划项目(No F15-199-1-28)
邹 丹(1971-),女,博士,副教授,研究方向:肿瘤调控,E-mail:3394984343@qq.com; 周伟强(1970-),男,博士,教授,研究方向:肿瘤调控,通讯作者,E-mail: zhouwq@hotmail.com
10.3969/j.issn.1001-1978.2017.03.006
A
1001-1978(2017)03-0317-05
R329.24;R392.11;R737.9;R977.1;R977.3
