近20a我国近海赤潮特点与发生规律*
2015-11-28丁德文林凤翱关春江
郭 皓,丁德文,林凤翱,关春江
(1.大连海事大学 环境科学与工程学院,辽宁 大连,116026;2.国家海洋环境监测中心,辽宁 大连,116023;3.近岸海域生态环境国家海洋局重点实验室,辽宁 大连,116023;4.国家海洋局 第一海洋研究所,山东 青岛,266061)
赤潮(Red-tide)是一种海洋生态异常现象,国际上将造成危害的赤潮称为有害藻华(Harmful Algal Blooms,HABs)[1]。近年来河口、内湾和沿岸水域污染不断加剧,水体富营养化程度日趋严重,赤潮发生的频率和危害程度明显上升,尤其是甲藻类赤潮有增多的趋势,但赤潮的发生机制仍未完全研究清楚[2]。
我国海域幅员辽阔,入海河流众多。随着海洋生态环境的改变,近年来我国海域赤潮灾害频繁发生,不仅影响到海洋生态系统和水产养殖业[3-5],因赤潮毒素引发的食品安全也对人类身体健康和生命安全带来了威胁[6]。我国赤潮记录起自20世纪30年代,但一直到80年代也仅是零星记录。随赤潮危害程度的增加,本世纪初开始,我国加大了对赤潮的监测与研究力度,国家海洋局在全国沿海省市设立了19个赤潮监控区,沿海监测部门在赤潮监控区内开展了高密度、高频率、多学科的综合调查监测,我国海域记录的赤潮事件明显增多,赤潮发生特点和规律逐渐显现,针对我国各海区[7-9]和不同海域[10-13]赤潮发生情况的研究也有陆续报道,但对我国沿海赤潮发生总体情况的研究较少[14-15]。本研究通过全面梳理和分析赤潮监测及研究数据,掌握了大量的一手资料[16-19]。因90年代之前数据记录的匮乏,本研究重点分析了从1995-2014年20a间我国近海赤潮发生的特点和规律,这对于掌握海区赤潮生物演替规律,揭示赤潮爆发机制,防范赤潮毒素危害等具有重要意义。
1 近20a我国近海赤潮总体概况
1995-2014年的20a间,我国近岸海域共记载赤潮事件约1 160次,累计发生面积约214 700km2。其中,造成较大经济损失的灾害性赤潮70余次,直接经济损失约36亿元。
如图1所示,1995-2000年,所记录的赤潮发生次数和累计面积相对较少,年均16次和4 250km2,但1998年和2000年因赤潮造成的直接经济损失分别为8.50亿元和2.73亿元;自2001年起,赤潮年发生次数维持在50次以上,年均次数、面积和损失分别为76次、13 520km2和1.72亿元。其中,2003年发生赤潮119次,为历年记录数量之最;2004,2005年连续2年赤潮累计面积均超过26 600km2;1998,2000,2010和2012年因赤潮灾害带来的直接经济损失均超过2.06亿元,其中,2012年米氏凯伦藻在我国东南沿海一带引发的赤潮对水产养殖业带来了毁灭性的打击,直接经济损失超过20亿元[20]。
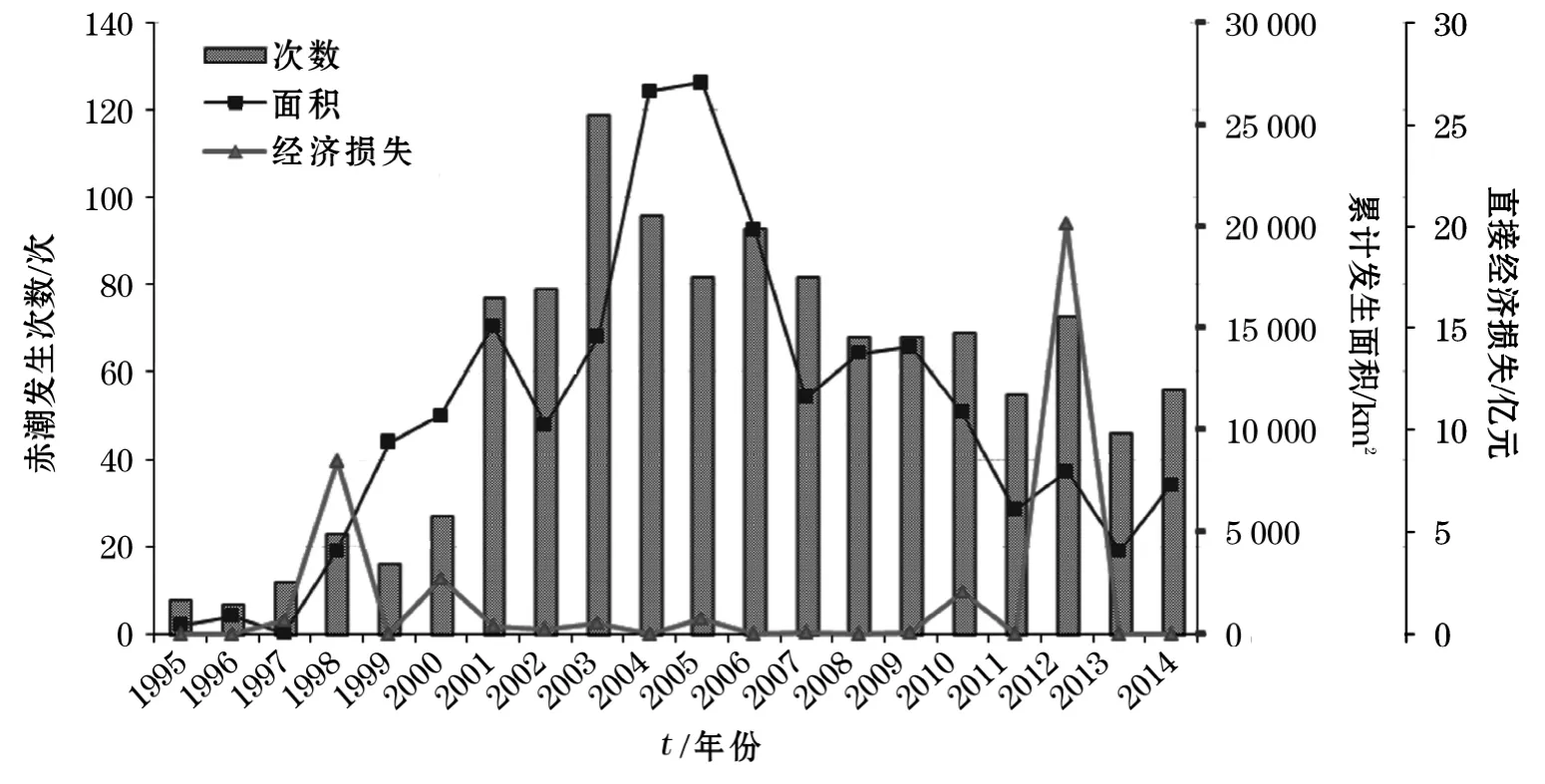
图1 1995-2014年我国沿海赤潮发生总体情况Fig.1 General situation of red tide in China during 1995-2014
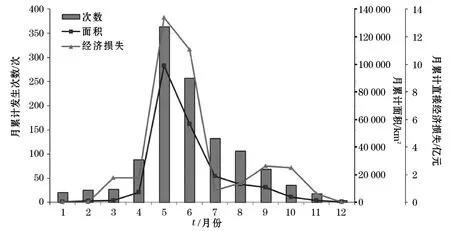
图2 1995-2014年我国沿海赤潮月发生情况Fig.2 Monthly situation of red tide in China during 1995-2014
我国海域发生的赤潮具有明显的季节特征。如图2所示,每年的4-9月份为赤潮高发季节,其发生次数、面积和经济损失呈现相同的变化趋势。其中,5,6,7月三个月份赤潮的发生次数、面积和经济损失占到全年赤潮总量的65.7%,81.4%和70.3%。每年冬季赤潮零星出现在我国南海海域,其发生范围和造成的损失也相对较小。
我国海域引发赤潮的生物门类主要包括甲藻门、硅藻门、着色鞭毛藻门、蓝藻门、绿藻门、棕鞭藻门和原生动物等,共90余种[17,21-22](图3)。其中,甲藻门、硅藻门和着色鞭毛藻门生物引发的赤潮分别占到50.0%,20.7%和7.5%,未定种类赤潮约占17.9%。主要赤潮生物为东海原甲藻(Prorocentrumdonghaiense)、夜光藻(Noctilucascintillans)、中肋骨条藻(Skeletonemacostatum)、米氏凯伦藻(Kareniamikimotoi),其发生次数和范围远高于其他种类;其次为红色中缢虫(Mesodiniumrubrum)、球形棕囊藻(Phaeocystisglobosa)、锥状斯氏藻(Scrippsiellatrochoidea)、海洋卡盾藻(Chattonellamarina)等种类。另外,一些有毒、有害藻类如叉角藻(Ceratiumfurca)、赤潮异弯藻(Heterosigmaakashiwo)、短凯伦藻(Karenia brevis)、多环旋沟藻(Cochlodiniumpolykrikoides)、多纹膝沟藻(Gonyaulaxpolygramma)、束毛藻(Trichodesmiumspp.)、红色赤潮藻(Akashiwosanguinea)、具刺膝沟藻(Gonyaulaxspinifera)、链状裸甲藻(Gymnodiniumcatenatum)、塔玛亚历山大藻(Alexandriumtamarense)、微小原甲藻(Prorocentrumminimum)等也时常引发赤潮。

图3 1995-2014年主要赤潮生物类群及其引发赤潮的次数Fig.3 Dominant red-tide groups and evoked cases during 1995-2014
不同类群赤潮生物引发的赤潮具有明显的季节性。如图4所示,硅藻类赤潮的高发期在每年的4-10月;甲藻类赤潮在每年的3月份开始旺发,夏、秋季节(4-10月份)进入高峰期,然后开始下降;着色鞭毛藻类赤潮全年表现较为平缓,没有明显的季节差别;绿藻-棕鞭藻类赤潮仅发生在春夏之交(4-6月份);蓝藻类赤潮高发期在每年的夏季(5-7月份)和秋冬季节;原生动物(红色中缢虫)赤潮全年均可发生。
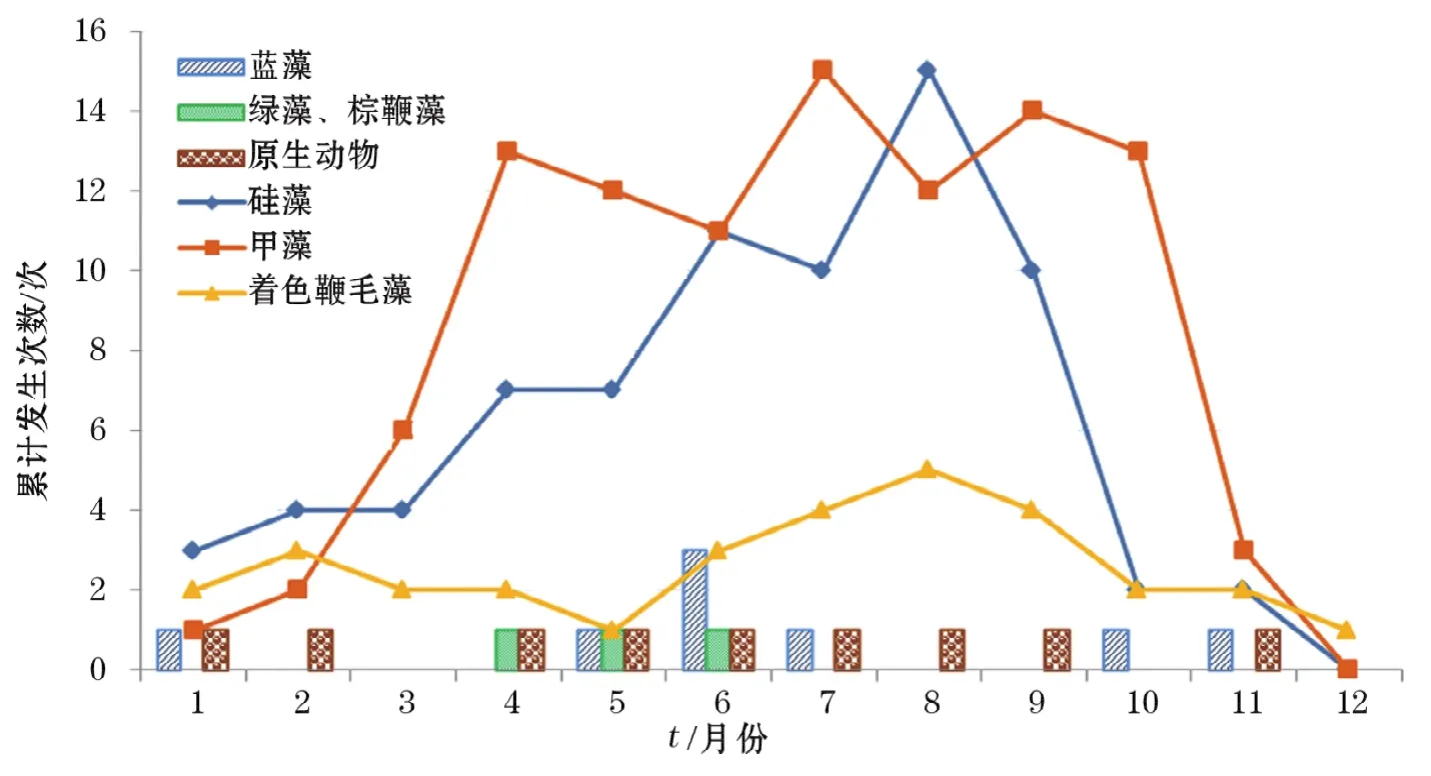
图4 不同月份主要赤潮生物类群及引发赤潮的次数Fig.4 Monthly change of primary red-tide groups and evoked cases
2 各海区赤潮发生特点分析
2.1 总体状况
我国沿海赤潮发生数量、影响范围和造成的经济损失具有显著的区域特点。东海海域为赤潮高发区和多发区,其发生次数、面积和损失分别占到赤潮总量的57.6%,62.4%和64.7%;其次为渤海,其数量、面积和损失分别占到赤潮总量的13.6%,23.4%和20.0%;黄海和南海海域相对较少(图5)。近20a发生造成较大直接经济损失的灾害性赤潮70余次,渤海、黄海、东海和南海海域分别占12.7%,7.0%,62.0%和18.3%。
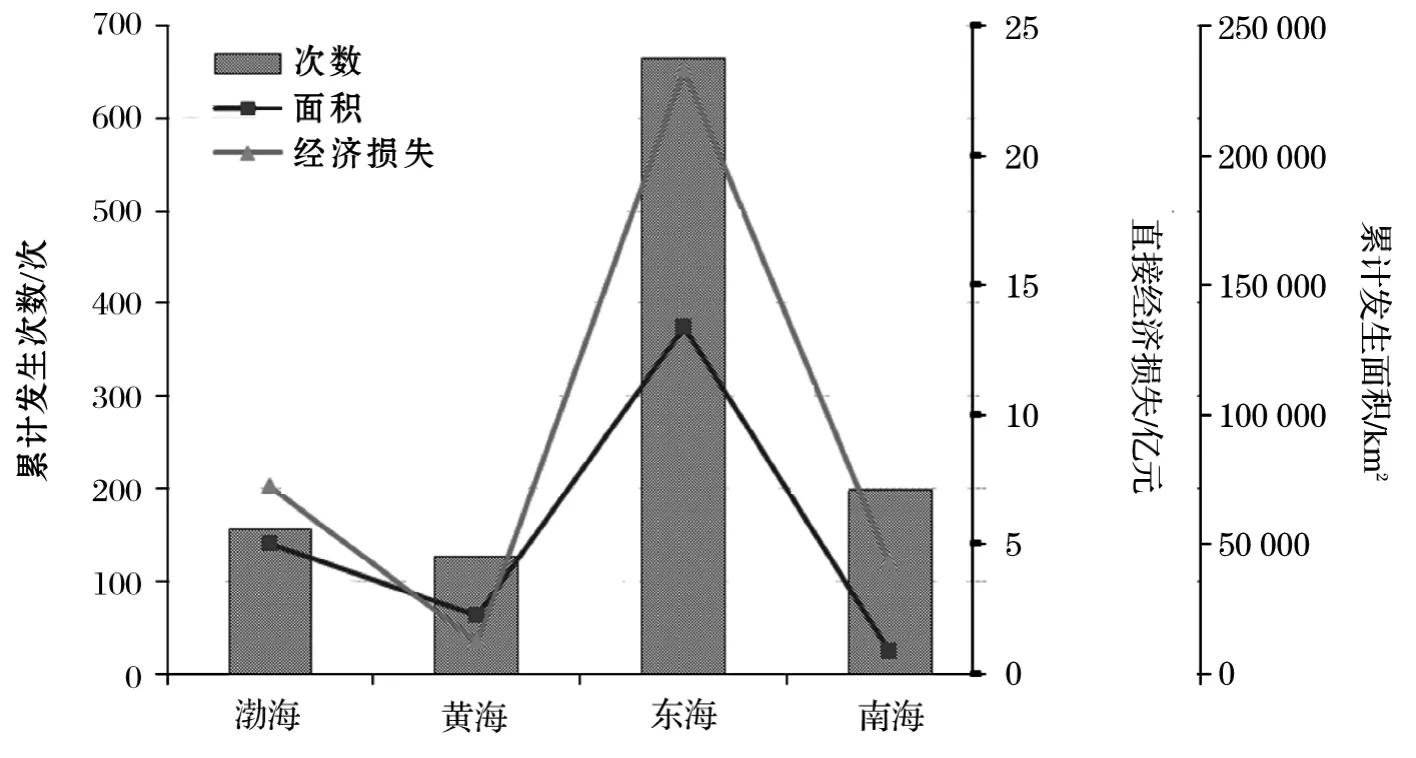
图5 1995-2014年各海区赤潮发生情况Fig.5 General situation of red tide in 4sea-areas during 1995-2014
2.2 年度变化
我国海域跨越暖温带、亚热带和热带三个气候带,环境差异较大,各海区赤潮发生情况各异(图6)。东海赤潮无论在数量、范围和损失方面都远高于其他3个海区,年平均次数33次,年均面积近6 700km2。其中,2003-2007年间年发生次数均超过50次,累计面积年均14 500km2,出现明显的峰值特征。
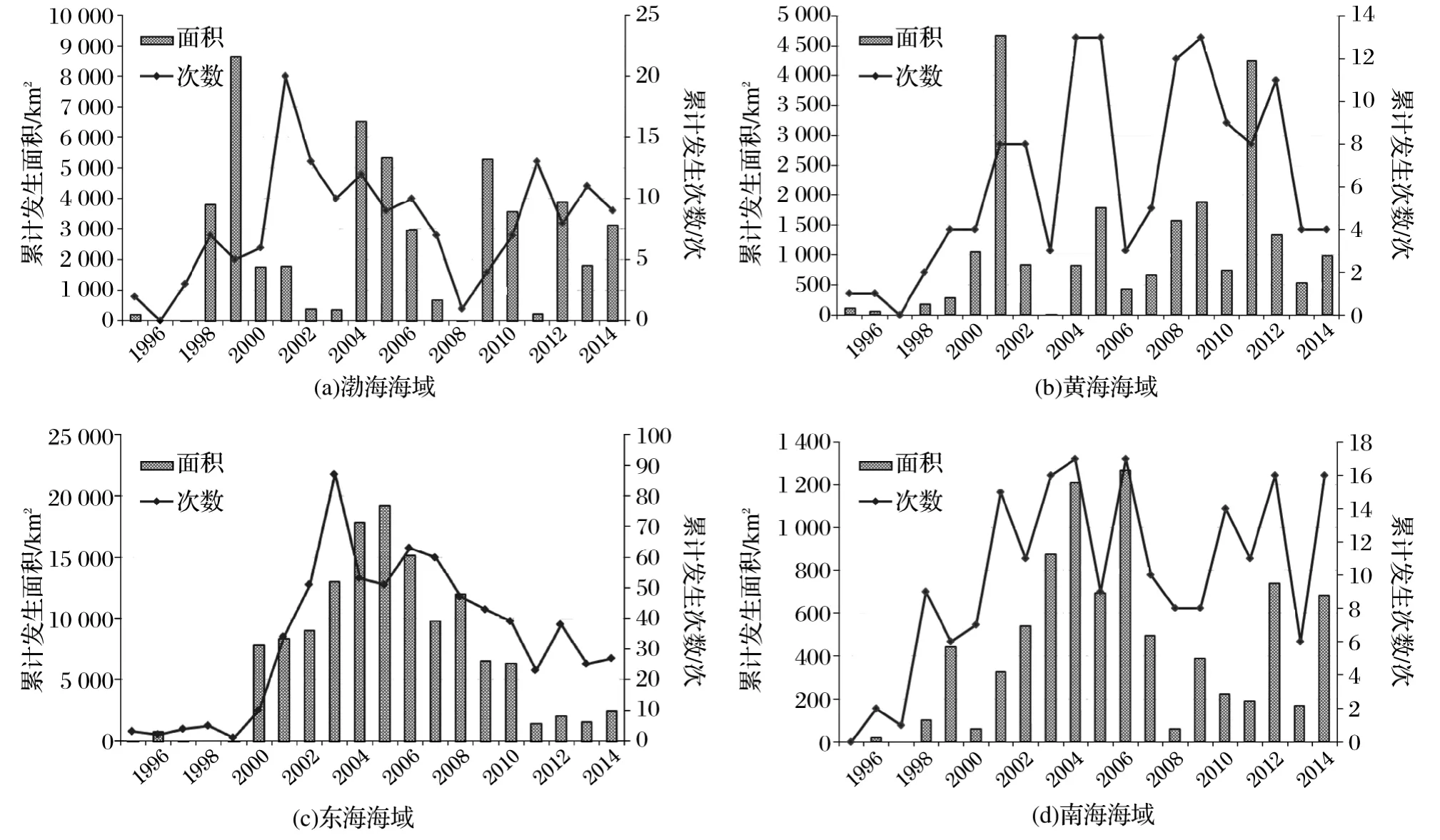
图6 1995-2014年各海区赤潮年度变化Fig.6 Annual change of red tide in 4sea-areas during 1995-2014
渤海为我国内海,赤潮发生次数虽略低于南海,但累计面积和造成的经济损失高于黄海和南海。其中,2001,2004-2006,2011-2013年赤潮发生次数较高,但最大累计面积出现在1999年,约8 640km2,其次为2004,2005和2009年,累计面积均超过5 200km2。
总体而言,黄海海域赤潮发生次数和范围相对较小,且年度变化较大,没有明显的规律性。其中,2004-2005,2008-2009及2012年赤潮发生次数较多,年均超过10次;面积较大的赤潮发生在2001和2011年,均超过4 200km2。
南海海域赤潮波动性相对平缓,高发年份在2001-2007,2010-2012年,年均10次左右;南海赤潮面积相对较小,年累计面积接近或超过500km2的赤潮出现在2002-2007年以及2012年和2014年。
2.3 季节变化
各海区赤潮发生显现出不同的季节特点(图7)。渤海赤潮高发季节为每年的5-7月,赤潮范围远大于其他月份;这3个月份赤潮数量和面积分别占到渤海全年赤潮总量的65.4%和80.1%;8-10月份,赤潮次数持续减少;进入秋冬季节后则极少发生。
黄海海域赤潮发生总趋势类似于渤海。每年的5-9月份为赤潮高发季节,其中8,9月份赤潮数量较高,而5月份赤潮累计面积达到最大;这5个月份赤潮的发生次数和面积分别占到全年赤潮总量的84.9%和94.8%。从10月份直到次年的4月份,黄海海域赤潮较少发生。
㉒王浦劬、赖先进:《中国公共政策扩散的模式与机制分析》,《北京大学学报》(哲学社会科学版)2013年第6期。
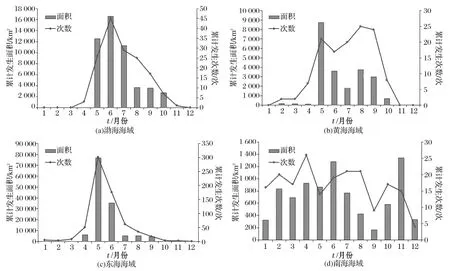
图7 1995-2014年不同月份各海区赤潮发生情况Fig.7 Monthly change of red tide in 4sea-areas during 1995-2014
东海海域赤潮具有显著的季节特征。进入4月,赤潮开始旺发,5-6月达到顶峰,赤潮数量和面积分别占到东海全年赤潮总量的71.8%和84.0%;随后其数量和面积直线下降。
南海海域赤潮季节变化不很明显,除9月份次数和面积均较少外,其余月份波动不大;赤潮面积在6月和11月各出现1个峰值,其他月份无显著差异。
2.4 主要赤潮生物类群
各海区引发赤潮的生物类群不尽相同(图8)。在渤海引发赤潮的主要生物类群包括甲藻、硅藻、着色鞭毛藻、棕鞭藻和原生动物等,近30种。其中,甲藻类、硅藻类和着色鞭毛藻赤潮数量分别占到渤海赤潮总量的48.4%,13.4%和13.4%;累计面积分别占到渤海赤潮总面积的45.0%,11.4%和17.3%。主要赤潮生物为夜光藻和抑食金球藻(Aureococcusanophagefferens),其中,夜光藻赤潮的数量和面积分别占到渤海赤潮总量的34.4%和29.5%;抑食金球藻赤潮从2009年开始在渤海秦皇岛沿岸海域连续出现[23-24],几年间累计面积近7 000km2,数量和面积占比分别达到到渤海赤潮总量的3.2%和14.0%。
黄海海域引发赤潮的主要生物类群包括甲藻、硅藻、着色鞭毛藻和原生动物等,其中,甲藻类、硅藻类和着色鞭毛藻赤潮数量分别占到渤海赤潮总量的40.5%,20.6%和13.5%;累计面积分别占到黄海赤潮总面积的36.9%,21.5%和7.9%。最主要的赤潮生物为夜光藻和中肋骨条藻,其中,夜光藻赤潮的数量和面积分别占到渤海赤潮总量的21.4%和30.9%;中肋骨条藻赤潮的数量和面积分别占到黄海赤潮总量的8.7%和9.3%。
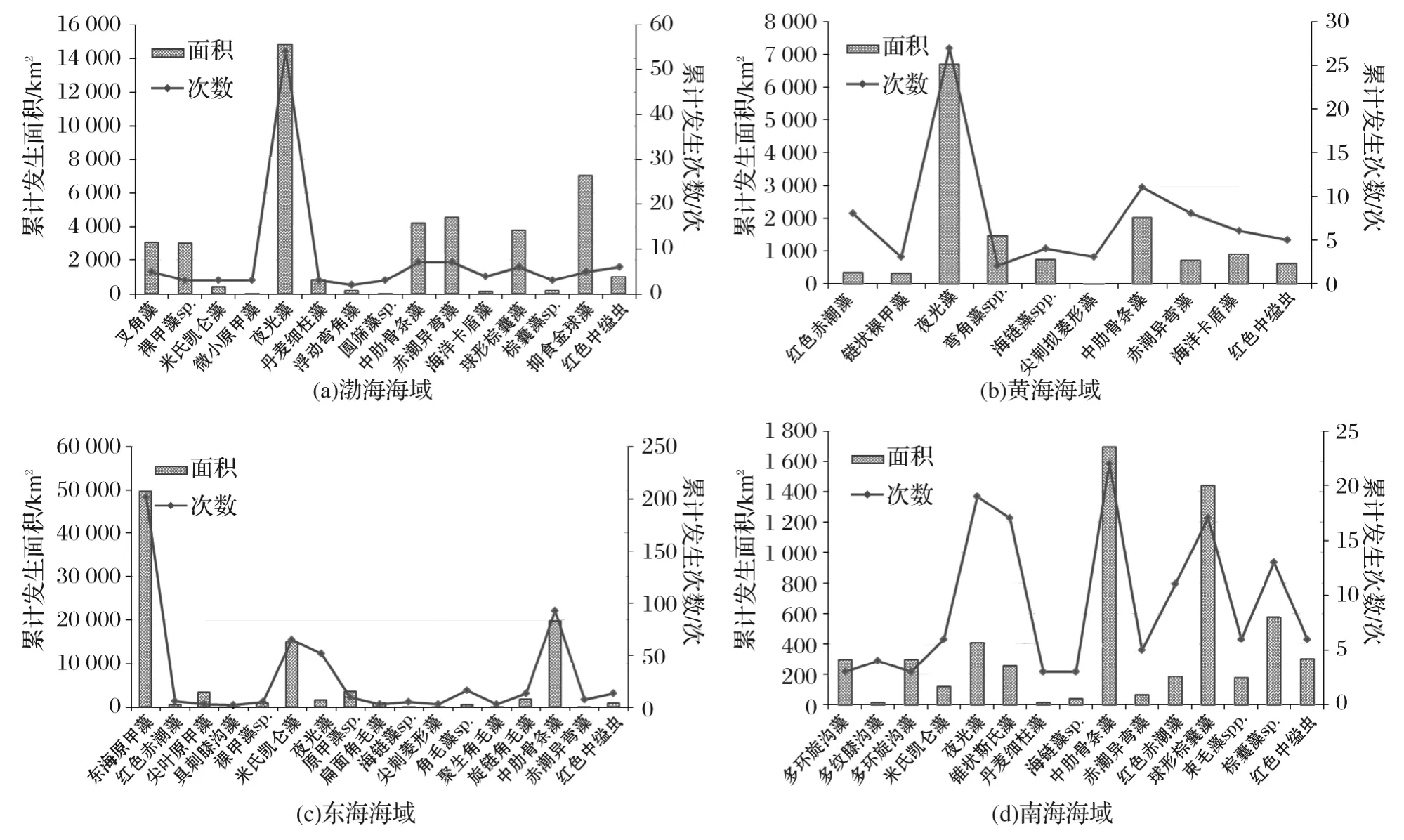
图8 各海区主要赤潮生物及其引发赤潮情况Fig.8 Main red-tide species and evoked cases in 4sea-areas
东海海域引发赤潮的主要生物类群包括甲藻、硅藻和原生动物等,其中,甲藻类、硅藻类赤潮数量各占到东海赤潮总量的44.0%;累计面积分别占到东海赤潮总面积的57.0%和19.1%。最主要的赤潮生物为东海原甲藻、中肋骨条藻、米氏凯伦藻和夜光藻,其中,东海原甲藻赤潮的数量和面积分别占到东海赤潮总量的30.2%和37.0%;中肋骨条藻赤潮的数量和面积分别占到东海赤潮总量的13.8%和14.6%;米氏凯伦藻赤潮的数量和面积分别占到东海赤潮总量的9.8%和11.2%。
南海海域引发赤潮的主要生物类群包括甲藻、硅藻和着色鞭毛藻等,其中,甲藻类、硅藻类和着色鞭毛藻赤潮数量分别占到南海赤潮总量的43.7%,18.1%和19.1%;累计面积分别占到南海赤潮总面积的24.5%,35.1%和24.8%。最主要的赤潮生物为中肋骨条藻、夜光藻和球形棕囊藻,其中,中肋骨条藻赤潮的数量和面积分别占到南海赤潮总量的11.1%和19.9%;夜光藻赤潮的数量和面积分别占到南海赤潮总量的9.5%和4.8%;球形棕囊藻赤潮的数量和面积分别占到南海赤潮总量的8.5%和16.9%。
2.5 灾害性赤潮及其造成的直接经济损失
本文所指的灾害性赤潮主要是对水产养殖业造成较大直接经济损失的赤潮事件,不包括对海洋生态系统、人类健康、旅游娱乐业等造成间接经济损失的藻华事件。1995-2014年,我国海域发生灾害性赤潮70余次,累计面积约31 400km2,仅分别占赤潮发生总数的6.1%和14.6%,但造成直接经济损失约36亿元。4个海区中,东海海域发生灾害性赤潮40余次,累计面积超过16 300km2,造成直接经济损失超过23亿元,分别占到赤潮总次数、总面积和总损失的62.0%,51.9%和64.7%;黄海海域发生灾害性赤潮的次数、面积和造成的经济损失相对较少,仅占7.0%,7.3%和3.5%;渤海和南海海域赤潮造成的经济损失各占20.0%和11.9%。
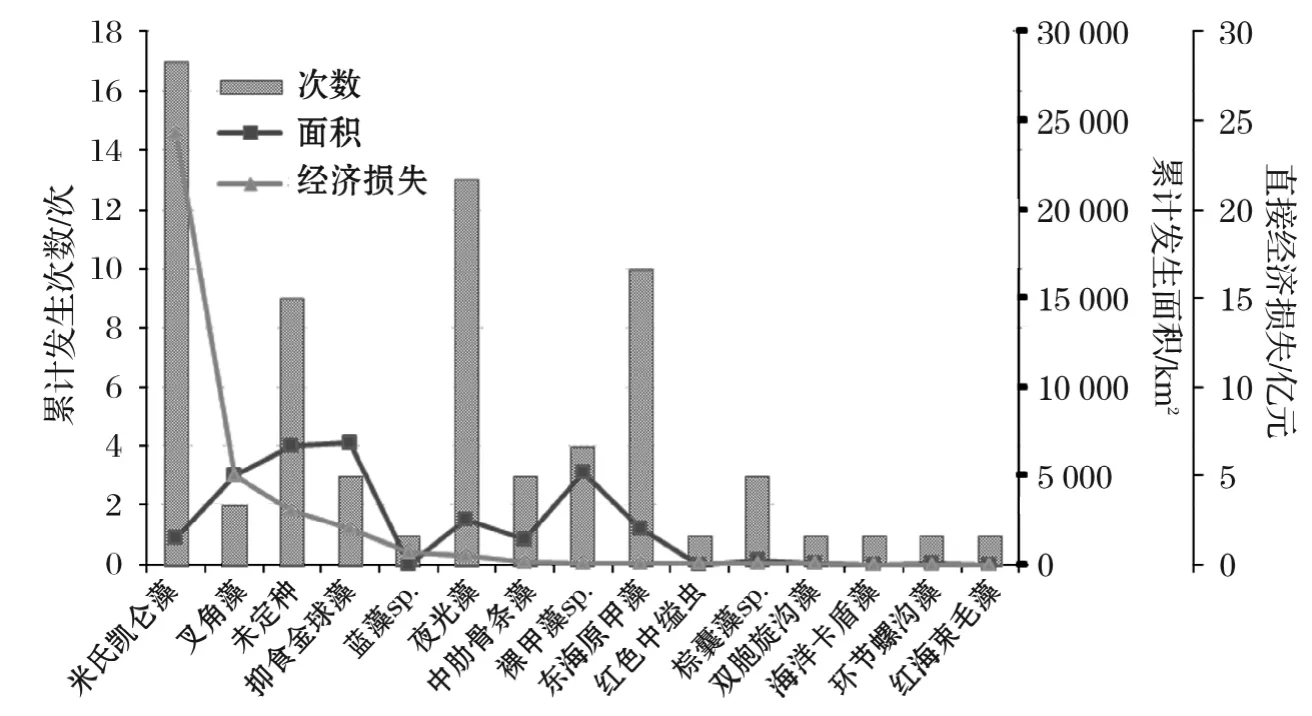
图9 1995-2014年我国沿海灾害性赤潮发生情况Fig.9 Disastrous red tides and direct economic losses during 1995-2014
统计数据表明(图9),20a间灾害性赤潮以甲藻类赤潮为主,其发生次数、面积和造成的经济损失分别占到灾害性赤潮总次数、总面积和总损失的67.6%,51.9%和83.6%;主要种类包括米氏凯伦藻[5,28]、叉角藻(Ceratiumfurca)、夜光藻、裸甲藻(Gymnodiniumsp.)、东海原甲藻、双胞旋沟藻(Cochlodiniumgeminatum,现更名为双胞多沟藻Polykrikosgeminatum)等[25-27]。其中,影响最大的为米氏凯伦藻赤潮[28],有17次造成较大经济损失,损失额度占到总量的67.5%;其次为叉角藻赤潮,损失额度占到总量的14.0%;夜光藻和东海原甲藻赤潮虽然发生次数较多,但造成的经济损失相对较少。着色鞭毛藻类赤潮也多次引发灾害性赤潮,其占比分别为11.3%,22.5%和5.7%,主要种类包括球形棕囊藻、棕囊藻sp.、海洋卡盾藻(Chattonellamarina)等。抑食金球藻赤潮具有明显的地域性和季节性,近几年主要发生在渤海海域,高发月份为5-7月,影响范围和造成的经济损失较大;红色中缢虫赤潮在我国海域全年均可发生,但造成的经济损失相对较少。
2.6 高发性赤潮及原因种分析
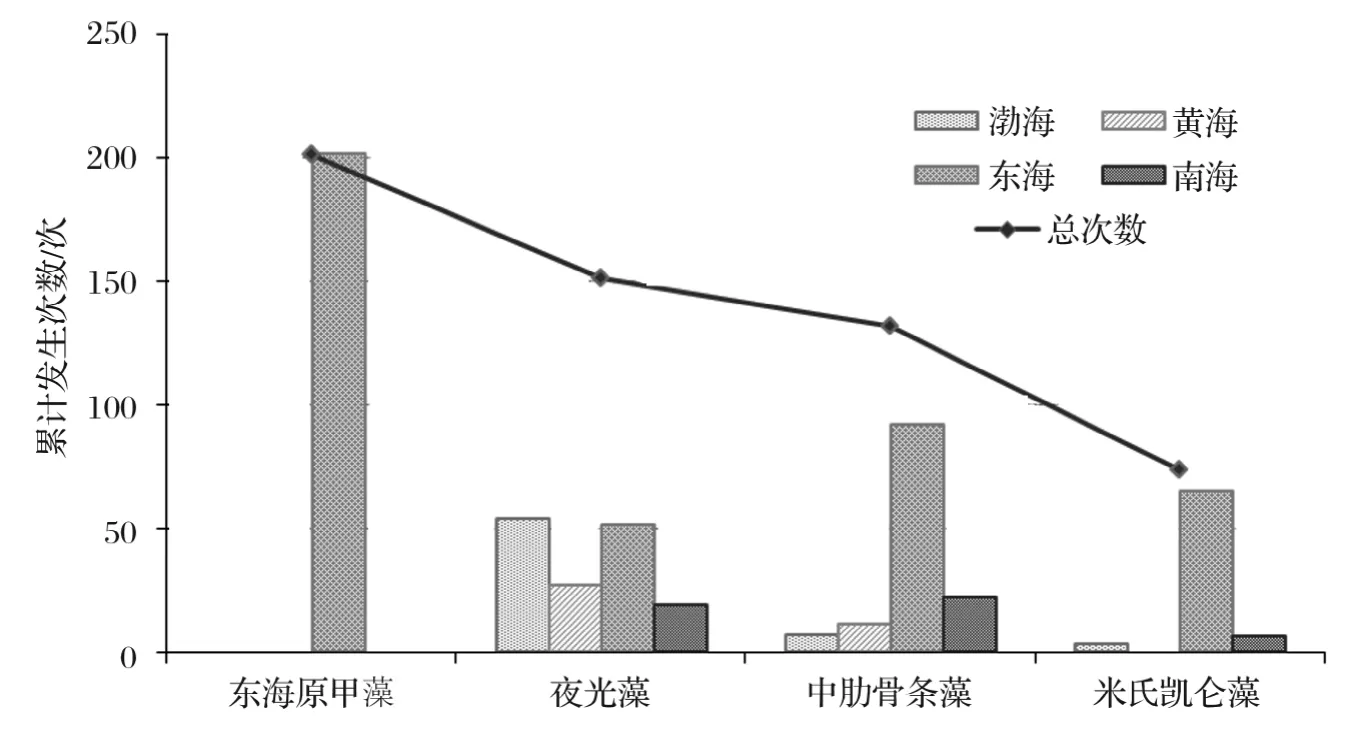
图10 各海区高发性赤潮数量及生物种类Fig.10 High incident red tides and species in 4sea areas
东海原甲藻主要分布在我国东海海域,其最适温度为18~22℃、盐度为22~31。自上世纪90年代后期,每年的4-9月,尤其是5,6月份东海海域都会爆发大规模的东海原甲藻赤潮。其中,2003年赤潮次数达32次。近20a间,共发生超过1 000km2的赤潮16次,累计面积约25 740km2,占东海原甲藻赤潮总面积的51.8%。因本种或其他伴生种引发的赤潮造成直接经济损失约910万元,主要危及养殖的鱼、虾、鲍鱼等。周名江和朱明远[29]研究显示,该种赤潮基本分布在27~31°N、沿30~50m等深线的海底地形陡变区海域,该海域大部分时间都处于富营养化状况。而在此之前,东海浮游植物群落保持着以硅藻为主、甲藻为次的结构特征,这可能与水温及营养盐组成变化有关。当水温低于20℃时,自然海域赤潮生物优势种为东海原甲藻并形成赤潮;水温超过23℃时,东海原甲藻种群迅速衰退,中肋骨条藻成为优势种群[37]。该区域溶解无机氮秋季以NO3-N为主,春季以NH4-N为主,东海原甲藻在自然海水中可利用的氮源及其广泛,高N/Si情况下甲藻的生长明显优于硅藻[30]。同时,N/P对东海原甲藻的生长有明显作用,研究表明,N/P<10时就会导致甲藻大量增殖,春季由台湾暖流带来的大量磷酸盐导致东海海域营养盐结构变化,这也是春季东海原甲藻赤潮爆发的一个重要原因[29-31]。
夜光藻在我国近海分布广泛,对环境的适应能力很强,在水温7.4~28.6℃,盐度14.8~32.9的范围内均能生长。研究结果显示,夜光藻大量繁殖的最适水温为18℃,适宜盐度为25~30,且对高盐(45)的耐受能力比对低盐(10)的耐受能力要高,表明夜光藻属于耐温耐盐上限较高的赤潮生物[32]。近20a该种在我国各海区几乎每年都会引发赤潮,其中,2003年就发生了23次夜光藻赤潮,累计面积达750km2。大面积赤潮主要发生在渤海和黄海海域,如1999-07,该种在辽东湾形成了6 330km2的赤潮;2005-06,在辽宁营口鲅鱼圈夜光藻赤潮面积达2 000km2;2011-05,鸭绿江口-辽宁大连长海海域赤潮面积达4 000km2,上述几次赤潮累计面积约占夜光藻赤潮总面积的58.8%。因本种引发的赤潮造成直接经济损失约5 340万元,主要受灾种类为鱼、虾、贝、藻类。夜光藻属于异养性甲藻,主要以浮游植物和浮游动物为饵料,在生产力较低的环境条件下难以形成数量巨大的种群。河口区域由于营养丰富,浮游植物生长迅速而改善了夜光藻生长的饵料基础,这可能是夜光藻赤潮爆发的原因之一[33]。食性实验研究结果显示,夜光藻对不同微藻的摄食强度依次为扁藻(Plalymonashelgolandica)>等鞭金藻(Isochrysisgalbana)>三角褐指藻(Phaeodactylum tricornyum);扁藻的密度与夜光藻生长呈明显相关关系。除摄食浮游植物外,夜光藻还摄食有机碎屑、小型浮游动物及其卵,摄食活动和营养繁殖具有昼夜节律性[33-35]。本种在赤潮爆发过程中易与中肋骨条藻交替成为优势种[36]。
中肋骨条藻为广温、广盐性的近岸性硅藻,是我国近海常见的赤潮生物,也是良好的污染指示生物,其增殖速率很快,细胞分裂次数达每天6次[36]。近20a间,共发生超过1 000km2的赤潮7次,累计面积约13 824km2,占中肋骨条藻赤潮总面积的50.2%。本种生存水温0~37℃、盐度13~36,最适水温24~28℃、盐度20~30[16]。史亚波通过培养实验,根据形态上和生理生态特征差异,把广布于我国近海的骨条藻种群分成中肋骨条S.costafum和热带骨条藻S.tropisum,前者具1~2个色素体,属于广温广盐种;后者具5~7个色素体,属于高温低盐种[32]。在营养充分、水文和气象条件相对稳定的情况下,中肋骨条藻在春、夏、秋三个季节均能形成繁殖高峰。特别是在长江下泄流量偏多,营养盐尤其是硅和氮的输入增加的情况下,更容易促使中肋骨条藻爆发性增殖并形成赤潮[36]。中肋骨条藻生长受N/P影响明显,主要是受到N限制。N限制和P限制培养对中肋骨条藻种群过程影响的差别在于,P限制下,细胞生物量(叶绿素a和细胞数量)最大值随温度的增加而增加;相反,在N限制下,细胞生物量最大值随温度的升高而降低。半连续培养下东海原甲藻和中肋骨条藻种群生长过程与种间竞争研究表明,东海赤潮高发区的营养盐浓度适合于东海原甲藻的种群生长,而低P限制可能成为中肋骨条藻等高营养盐需求藻类生长的限制因子[37]。
米氏凯伦藻是一种典型的有毒赤潮藻类,在澳大利亚、日本、南美、北非和我国均引发过赤潮并造成了严重的经济损失。20世纪初,我国沿海频繁爆发米氏凯伦藻赤潮并由南向北扩展[28]。近20a由该种引发的赤潮近80次,累计面积约15 500km2。其中,1998-03珠江口爆发的米氏凯伦藻赤潮给粤港两地造成经济损失高达3.5亿元,2012年,在浙江、福建多个海区连续爆发米氏凯伦藻赤潮,造成大量养殖鲍死亡,直接经济损失超过20亿元。米氏凯伦藻适温范围较窄,20~24℃生长良好。对盐度变化敏感,适宜盐度27.9~30.5,剧烈的盐度变化导致渗透压的改变,容易引起细胞破裂。本种有明显的昼夜垂直移动,主要在海表至海表下2m间的水层活动[5]。米氏凯伦藻赤潮的发生与降雨相关性明显,除降雨带来丰富的营养盐和微量元素外,低光照对米氏凯伦藻赤潮的形成也具有一定作用。本种对营养盐要求较高,研究表明,高营养指数和磷酸盐是米氏凯伦藻形成赤潮的重要条件,微量元素也具有较高的促进作用[28]。米氏凯伦藻赤潮的发生往往引发海洋生物的大量死亡并造成严重的经济损失,其产生的溶血毒素和鱼毒素,可溶解鱼类的鳃组织引起生物死亡;米氏凯伦藻赤潮对海洋生态环境也产生显著影响,如直接接触活的米氏凯伦藻细胞可以导致浮游动物死亡[5]。近20a来,因米氏凯伦藻赤潮造成的直接经济损失约24.3亿元,占到赤潮灾害总损失的67.5%,主要受灾种类为养殖鲍鱼和其他水产品种类。
3 结论
近年来我国海域赤潮发生特点是:赤潮高发期在气象学上的春季和夏季;赤潮发生频率增多、持续时间加长、危害程度增大;主要赤潮生物种类尤其是有毒赤潮藻类增多;赤潮频发海域多为受无机氮和磷酸盐污染较重的海域;大面积赤潮主要集中在近岸、近海和河口附近海域。
1)我国近岸海域全年均可发生赤潮,但随海区不同,赤潮高发期有所差别。南海海域赤潮季节变化不显著,其他3个海区的赤潮高发期集中在每年的5-9月份。
2)我国近岸海域均有赤潮发生记录,高发区域在渤海和东南沿海;大面积赤潮也主要集中在渤海和东海近岸海域。其中,东海赤潮在数量、范围和损失方面远高于其他3个海区,并且具有明显的季节特征。
3)我国海域引发赤潮的主要生物类群为甲藻、硅藻、着色鞭毛藻、蓝藻和原生动物等,其中,有毒赤潮生物20余种;甲藻类赤潮占到赤潮总数的一半;引发赤潮次数最多的为东海原甲藻、夜光藻、中肋骨条和米氏凯伦藻等。
4)灾害性赤潮以甲藻类赤潮为主,其中,米氏凯伦藻、叉角藻赤潮造成巨大经济损失;主要受灾品种为养殖贝类、鱼类和虾蟹类,海洋生态环境也受到严重影响。
5)甲藻类赤潮发生次数明显高于其他种类,说明环境条件在赤潮优势种的演替过程中起到了重要作用。
6)今后的工作中应更加关注有害藻华生物,结合长时空序列数据进行深入分析,这有助于我们掌握赤潮生物演替规律,对赤潮爆发作出预测预警,及时防范赤潮风险。
(References):
[1]HALLEGRAEFF G M,ANDERBELLA D M,CEMBELLA A D.Manual on Harmful Marine Microalgae[M].Paris:UNESCO,1995.
[2]HUANG L M,HUANG X P,SONG X Y,et al.Frequent occurrence areas of red tide and its ecological characteristics in Chinese coastal waters[J].Ecologic Science,2003,22(3):252-256.黄良民,黄小平,宋星宇,等.我国近海赤潮多发区域及其生态学特征[J].生态科学,2003,22(3):252-256.
[3]CHEN Q Z,HE D H.An approach of red tide of dinoflagellates impacts on abalone aquaculture in Fujian Province and countermeasures[J].Journal of Fisheries of China.2000,24(2):151-156.陈全震,何德华.甲藻赤潮对养鲍业的危害及其防治探讨[J].水产学报,2000,24(2):151-156.
[4]CHEN S,TONG M M,JIANG T J,et al.The assessment of aquaculture loss caused by red tides[J].Journal of Fisheries of China,2009,33(4):610-616.陈舜,佟蒙蒙,江天久,等.赤潮灾害对水产养殖业损失的分级评估[J].水产学报,2009,33(4):610-616.
[5]CHNE B H,XIE E Y,GAO Y H,et al.Toxic effects of red tide caused byKareniamikimotoion marine organisms[J].Journal of Fujian Fisheries,2015,37(3):241-250.陈宝红,谢尔艺,高亚辉,等.米氏凯伦藻对海洋生物致毒作用的研究进展[J].福建水产,2015,37(3):241-250.
[6]LIN Y T,JIA X P,YANG M L,et al.Paralytic shellfish posion in seashells collected along the coast of China and its origin[J].Journal of Fisheries of China,2001,25(5):479-481.林燕棠,贾晓平,杨美兰,等.我国海产贝类体中的麻痹性毒素及其来源[J].水产学报,2001,25(5):479-481.
[7]LIN F A,LU X W,LUO H,et al.History,status and characteristics of red tide in Bohai Sea[J].Marine Environmental Science,2008,27(supp.2):1-5.林凤翱,卢兴旺,洛昊,等.渤海赤潮的历史、现状及其特点[J].海洋环境科学,2008,27(增刊2):1-5.
[8]YE S F,HUANG X Q.HABs in East China Sea:Surveillance and monitoring[J].Marine Environmental Science,2003,22(2):11-14.叶属峰,黄秀清.东海赤潮及其监视监测[J].海洋环境科学,2003,22(2):11-14.
[9]WANG S F,TANG D L.Analysis of harmful algal blooms associated with ocean environments in the South China Sea[J].Marine Science Bulletin,2010,29(5):577-583.王素芬,唐丹玲.南海赤潮的时空分布特征及其与南海环境关系的综合分析[J].海洋通报,2010,29(5):577-583.
[10]WANG H Q,ZHANG Y B.Investigation of red tide organisms and their regularities in Dalian Bay[J].Marine Environmental Science,1992,11(2):28-33.王惠卿,张永波.大连湾赤潮生物及其规律的探讨[J].海洋环境科学,1992,11(2):28-33.
[11]XIA P,LU D D,ZHU D D,et al.Trend and characteristics of harmful algal blooms in Zhejiang coastal waters[J].Journal of Marine Sciences,2007,25(2):47-56.夏平,陆斗定,朱德弟,等.浙江近岸海域赤潮发生的趋势与特点[J].海洋学研究,2007,25(2):47-56.
[12]LI X D.Analysis on characteristics of red tide in Fujian coastal waters during the last 10years[J].Environmental Science,2012,33(7):2210-2216.李雪丁,福建沿海近10a赤潮基本特征分析[J].环境科学,2012,33(7):2210-2216.
[13]QIAN H L,LIANG S.QI Y Z.Study of the characteristics and the causes of formation on the red tides in coastal Guangdong Sea[J].Ecologic Science,2000,19(3):8-16.钱宏林,梁松,齐雨藻.广东沿海赤潮的特点及成因研究[J].生态科学,2000,19(3):8-16.
[14]LIANG S,QIAN H L,QI Y Z.Problem on the red tide in coastal China Sea[J].Ecologic Science,2000,19(4):44-50.梁松,钱宏林,齐雨藻.中国沿海的赤潮问题[J].生态科学,2000,19(4):44-50.
[15]LUO H,MA M H,LIANG B,et al.Basic characteristics and mitigation countermeasures of red tides in China seas[J].Marine Science Bulletin,2013,32(5):595-600.洛昊,马明辉,梁斌,等.中国近海赤潮基本特征与减灾对策[J].海洋通报,2013,32(5):595-600.
[16]QI Y Z,et al.Red tide in Chinese coastal waters[M].Beijing:Science Press,2004.齐雨藻,等.中国沿海赤潮[M].北京:科学出版社,2003.
[17]GUO H.Illustrations of planktons responsible for the blooms in Chinese coastal waters[M].Beijing:Science Press,2004.郭皓.中国近海赤潮生物图谱[M].北京:海洋出版社,2004.
[18]State Oceanic Administration.Bulletin of marine environmental status of China[EB/OL].(2015-03-11)[2015-09-06].http://www.soa.gov.cn.国家海洋局.中国海洋环境状况公报[EB/OL].(2015-03-11)[2015-09-06].http://www.soa.gov.cn.
[19]State Oceanic Administration,Bulletin of Marine Disaster of China[EB/OL].(2015-03-03)[2015-09-06].http://www.soa.gov.cn.国家海洋局.中国海洋灾害公报[EB/OL].(2015-03-11)[2015-09-06].http://www.soa.gov.cn.
[20]ZHOU M J,ZHU M Y,ZHANG J.Status of harmful algal blooms and related research activities in China[J].Chinesr Bulletin of Life Sciences,2001,13(2):54-59.周名江,朱明远,张经.中国赤潮的发生趋势和研究进展[J].生命科学,2001,13(2):54-59.
[21]ONG,HAN C,WILHELM,STEVEN W,et al.Analyses of the complete chloroplast genome sequences of two members of the pelagophyceae:Aureococcus Anophagefferens CCMP 1984andAureoumbraLagunensisCCMP 15071[J].Journal of Phycology,2010,46(3):602-615.
[22]CHEN Y H,LIANG J R,CHEN C P,et al.Brown tide:A new ecosystem disruptive algal bloom[J].Chinese Journal of Ecology,2015,34(1):274-281.陈杨航,梁君荣,陈长平等.褐潮——一种新型生态系统破坏性藻华[J].生态学杂志,2015,34(1):274-281.
[23]ZHNAG Q C,QIU L M,YU R C,et al.Emergence of brown tides caused byAureococcusanophagefferensHargraves et Sieburth in China[J].Harmful Algae,2012,19:117-124.
[24]GU B,ZHEN Y,MI T Z.Characterization of phytoplankton community in the coastal waters of Qinghuangdao during brown tide[J].Periodical of Ocean University of China,2015,45(7):064-072.古彬,甄毓,米铁柱.秦皇岛扇贝养殖区褐潮暴发期浮游微藻群落结构特点研究[J].中国海洋大学学报,2015,45(7):064-072.
[25]WANG Z H,LIN L C,KANG W,et al.Progresses in studies on harmful algal bloom speciesCochlodiniumSchutt[J].Journal of Jinan University(Natural Science),2011,32(1):110-114.王朝晖,林朗聪,康伟,等.有毒有害赤潮藻类旋沟藻(Cochlodinium Schutt)研究进展[J].暨南大学学报(自然科学版),2011,32(1):110-114.
[26]OU L J,ZHANG Y Y,LI Y,et al.The outbreak ofCochlodiniumgeminatumbloom in Zhuhai,Guangdong[J].Journal of Tropical O-ceanography.2010,29(1):57-61.林坚,张玉宇,李扬,等.广东珠海双胞旋沟藻Cochlodinium geminatum赤潮事件分析[J].热带海洋学报,2010,29(1):57-61.
[27]QIU D,HUANG L,LIU S,et al.Apical groove type and molecular phylogeny suggests reclassification ofCochlodiniumgeminatumasPolykrikosgeminatum[J].PLoS ONE 2013,8(8):e71346.
[28]LONG H,DU Q.Primary research onKareniamikimotoibloom in Fujian Coast[J].Journal of Fujian Fisheries,2005,(4):22-26.龙华,杜琦.福建沿海米氏凯伦藻赤潮的初步研究[J].2005,(4):22-26.
[29]ZHOU M J,ZHU M Y,Progress of the Project Ecology and oceanography of harmful algal blooms in China[J].Advances in Earth Science,2006,21(7):673-679.周名江,朱明远.我国近海有害赤潮发生的生态学、海洋学机制及预测防治研究进展[J].地球科学进展,2006,21(7):673-679.
[30]ZHANG C S.The characteristic and effects of nutrient during the process of HAB in Changjiang River Estuary and its adjacent area[D].Qingdao:Ocean University of China,2008.张传松,长江口及邻近海域赤潮生消过程特征及其营养盐效应分析[D].青岛:中国海洋大学,2008.
[31]XU Z L.The inter-annual variations inNoctilucascintillansabundance and eutrophication Changjiang estuary[J].Oceanologia et Limnologia Sinica,2009,40(6):793-797.徐兆礼.长江口夜光藻(Noctilucascintillans)年间变化和水域富营养化趋势[J].海洋与湖沼,2009,40(6):793-797.
[32]ZHAO D Z,CHENG J L,GUO H,et al.The study on the regularity of red-tide in Huang-Bo Hai Sea[M]∥ZHAO D Z.In Research corpus of monitoring and assessment of red tide hazards in Bohai Sea.Beijing:Ocean Press,2000,52-66.赵冬至,陈江麟,郭皓,等.渤海、黄海赤潮发生规律研究[M]∥赵冬至.渤海赤潮灾害监测与评估研究文集.北京:海洋出版社,2000,52-66.
[33]ZHOU C X,WU Y L,ZOU J Z.Nutrient dynamics ofNoctilucascintillans(Macartney)[J].Oceanologia et Limnologia Sinica,1994,55(2):152-157.周成旭,吴玉霖,邹景忠.夜光藻的营养动力学[J].海洋与湖沼,1994,55(2):152-157.
[34]WU Y L,ZHOU C X,ZHANG Y S.Laboratory culture ofNoctilucascintillans(Macartney)[J].Oceanologia et Limnologia Sinica,1994,25(2):165-167.吴玉霖,周成旭,张永山.夜光藻的室内培养[J].海洋与湖沼,1994,25(2):165-167.
[35]CHEN H H,QI S.The feeding and vegetative reproduction diurnal rhythms ofNoctilucascintillans[J].Journal of Jinan University,1991,12(3):104-111.陈汉辉,杞桑.夜光藻摄食和营养繁殖的昼夜节律[J].暨南大学学报,1991,12(3):104-111.
[36]Lu D D,GOBEL J,WANG C S,et al.Monitoring of harmful microalgae and nowcasting of red tides in Zhejiang coastal water[J].Donghai Marine Science,2000,18(2):33-44.陆斗定,J.Gobel,王春生,等.浙江海区赤潮生物监测与赤潮实时预测[J].东海海洋,2000,18(2):33-44.
[37]WANG Z L,LI R X,ZH U M Y,et al.Study on population growth processes and interspecific competition ofProrocentrumdonghaienseandSkeletonemacostatumin semi-continuous dilution experiments[J].Advances in Marine Science,2006,24(4):495-503.王宗灵,李瑞香,朱明远,等.半连续培养下东海原甲藻和中肋骨条藻种群生长过程与种间竞争研究[J].海洋科学进展,2006,24(4):495-503.
