不同氮磷比对福建沿海米氏凯伦藻生长的影响
2017-08-30潘非斐
潘非斐, 吴 楠
(福建省海洋环境与渔业资源监测中心,福建 福州 350003)
不同氮磷比对福建沿海米氏凯伦藻生长的影响
潘非斐, 吴 楠
(福建省海洋环境与渔业资源监测中心,福建 福州 350003)
本文在不同氮磷比(N∶P=4∶1、8∶1、16∶1、32∶1、80∶1)培养条件下,对福建沿海赤潮海域分离的米氏凯伦藻(Kareniamikimotoi)进行培养,研究其生长特性。实验结果表明:不同的氮磷比对米氏凯伦藻的生长有明显的影响。过高或过低的氮磷比均不适合米氏凯伦藻的生长,该藻在N∶P=32∶1条件下比生长率最快,为0.33 d-1。米氏凯伦藻对氮的需求高于磷,在适当的磷限制环境中能够维持更长的生长周期。引发赤潮的主要原因并不是由于米氏凯伦藻赤潮暴发海域的低氮磷比,而是赤潮暴发过程中,米氏凯伦藻对营养盐的大量消耗,尤其是对氮的消耗。
氮磷比;赤潮;米氏凯伦藻
米氏凯伦藻(Kareniamikimotoi),属于裸甲藻目(Gymnodiniales)、凯伦藻属(Karenia),是一种典型的有毒有害赤潮种。近年来,该种在我国沿海频繁引发赤潮,造成了严重的渔业经济损失。2012年5—6月,福建福鼎、霞浦、黄岐、平潭、湄洲岛等10处海域先后发生米氏凯伦藻赤潮,持续时间达20 d,累计面积323 km2,并导致水产养殖贝类尤其是养殖鲍的大规模死亡,直接经济损失高达20.11亿元[1]。
氮和磷是浮游植物生长的重要营养物质,通常浮游植物是按照Redfield系数(N/P=16)吸收利用海水中的氮磷,当海水中的营养盐摩尔比值偏离该系数时,则会对浮游植物的生长产生一定的限制作用,因此,海水中的氮磷比将影响海洋浮游植物的生长速率和细胞内物质合成[2]。但是Redfield系数并不适合所有浮游植物,不同的浮游植物有适合其自生长的最适氮磷比[3-4]。因此,研究米氏凯伦藻赤潮发生时的氮磷浓度对藻类生长的影响,对探究赤潮发生规律、演替和成因,并进行赤潮预测预报,减少对水产养殖业的损害有着重要的意义。本文结合2012年福建省沿海米氏凯伦藻赤潮发生时的现场环境条件,研究了不同氮磷浓度比对米氏凯伦藻生长的影响,以期为福建沿海米氏凯伦藻赤潮监测预警提供理论基础。
1 材料与方法
1.1 藻种
米氏凯伦藻藻种来源于2012年福建米氏凯伦藻赤潮发生海域。实验前取200 mL处于对数期的米氏凯伦藻(密度达104个/mL以上)离心,培养于无氮无磷的f/2培养基,培养4 d后待用。
1.2 培养条件
实验所用的器皿均用5% HCl浸泡后,以蒸馏水反复冲洗,灭菌后备用。培养基采用天然外海水通过0.45 μm醋酸纤维膜过滤,按照不加硅酸盐的f/2配方配制,氮、磷浓度按实验要求添加,经120℃高温蒸汽灭菌15 min后备用。培养温度为(20±2)℃,盐度利用灭菌蒸馏水稀释至盐度约为28.4,光照强度为2 500 Lx,光暗比为12 h∶12 h。
1.3 实验方法
以NO3-N为氮源、NaH2PO4·2H2O为磷源,通过固定磷浓度,改变氮浓度梯度设置初始氮磷比进行实验。设置初始磷浓度为0.8 μmol/L(约0.024 8 mg/L,参考2012年连江黄岐米氏凯伦藻赤潮发生期间磷浓度含量),初始氮浓度依次为0(对照组,即不添加氮元素)、3.2、6.4、12.8、25.6、64.0 μmol/L,使得氮磷比依次为0、4、8、16、32、80。每个浓度梯度设置两个平行样,每个平行样培养体积为600 mL,接种的初始密度为5×102个/mL。
细胞计数:自接种日起,每天在上午10∶30左右取样,每个梯度取2个样,每个样取1 mL藻液,用鲁哥试剂固定,用0.1 mL的浮游生物计数框,在BX51生物显微镜下进行细胞计数,实验共观察15 d。
N、P测定:与细胞计数同步进行,取20 mL藻液,用0.45 μm混合纤维素酯滤膜过滤后,4℃保存,用德国Brant Luebbe GmbH AA3HR型流动化学分析仪测定。
1.4 数据处理
细胞比生长率按以下公式计算:μ=(lnC1-lnC2)/(T1-T2),其中C1和C2分别是藻类在T1和T2时刻的细胞数量。
利用SPSS18.0软件在单因素方差分析(one-way ANOVA)的基础上,采用LSD多重比较法检验不同氮磷比处理间的差异(P<0.05)。
2 结果
2.1 不同氮磷比条件下米氏凯伦藻的生长情况
在不同初始氮磷比条件下米氏凯伦藻的生长特征如表1和图1所示,各组米氏凯伦藻细胞数均能达到103个/mL。最高细胞数在N/P为0~16之间是随着氮磷比的增加而增加,在N/P =32时则略有下降,低于N/P =16;但在N/P =80时最高细胞数则大幅上升,达到3.74×103个/mL,远高于其他组;最大比生长率在N/P为0~32之间的变化趋势也是随着氮磷比的增加而增加,最高值出现在N/P =32条件下,达到0.33 d-1,而在N/P =80时最大比生长率却大幅下降,仅为0.20 d-1。

表1 不同氮磷比下米氏凯伦藻的最高细胞数和最大比生长率

从图1可以看出,各组米氏凯伦藻的生长在前三天均处于停滞期,生长缓慢,差异不大;但从第4天开始,受不同的氮磷比影响,米氏凯伦藻的生长趋势在不同的培养液中明显不同。当N/P =4时,米氏凯伦藻细胞数量虽有增加,但生长缓慢,在第6天达到细胞密度最高值,仅为1.31×103个/mL,第7天细胞数量迅速下降,提前进入衰亡期,期间并未经过平稳期,与对照组相比未有显著差异(P>0.05)。N/P=8与对照组相比具有极显著差异(P<0.01),藻类在停滞期后快速生长,经历了3 d的对数期,第6天达到生长高峰,最高细胞数达到2.66×103个/mL,但随后第7天细胞密度迅速下降,提前进入衰亡期。N/P=16与对照组之间也存在极显著差异(P<0.01),在第4天进入对数期,第7天达到生长高峰,最高细胞数为3.01×103个/mL,随后也是提前进入衰亡期。N/P=32和N/P=80也分别与对照组间存在极显著差异(P<0.01),二者由于前期受到高氮浓度的抑制作用,生长缓慢,分别在第5天和第6天进入对数期,随后细胞快速生长,在第7天和第11天达到细胞最大值,分别为2.68×103个/mL和3.74×103个/mL;与前几组不同的是,在N/P=32和N/P=80条件下的米氏凯伦藻在达到细胞最大值后并未直接进入衰亡期,而是进入平稳期,直至实验结束仍维持较高的细胞密度。多重比较分析结果显示,实验期间除了N/P=16条件下米氏凯伦藻细胞密度均值与N/P=8及N/P=32的差异并不显著(P>0.05),以及在N/P=80条件下米氏凯伦藻细胞密度均值与N/P=32的差异亦不显著(P>0.05)外,其余各组间均存在显著差异(P<0.05)。
2.2 不同氮磷比下米氏凯伦藻对氮的消耗特征
从图2中可以看出,除N/P=4组外,其余各组在实验条件下均是于第2天氮浓度有所下降,而第3天则与第2天持平,随即第4天各组氮浓度均开始持续下降。在N/P=4条件下,氮浓度的消耗主要集中在前4天,第5天氮浓度开始迅速下降,至第15天实验结束时,培养液中氮的含量减少了3.275 μmol/L,氮的吸收率为99.46%。在N/P=8条件下,氮浓度从第4天开始大幅下降,至第15天实验结束时,培养液中氮的含量接近于0,共减少了9.389 μmol/L,氮的吸收率为99.70%。当N/P=16时,第2天和第4天氮的浓度均有大幅度下降,随后持续平缓下降,直至第15天实验结束时,培养液中氮的含量减少了18.004 μmol/L,吸收率为99.72%。当N/P=32时,氮浓度在第4天开始持续下降,至第15天实验结束时,培养液中氮的含量减少了29.300 μmol/L,吸收率为94.79%。在N/P=80条件下,氮消耗分为2个阶段,从初始到第12天为氮的快速消耗阶段,氮含量消耗至29.071 μmol/L;从第13天开始进入氮的缓慢消耗阶段,氮略有消耗,但消耗量不大;此后至第15天实验结束,培养液中的氮浓度虽然大量减少,但是与其它组氮浓度接近于0的情况相比较,该条件下的氮浓度仍然较高,减少了51.746 μmol/L,氮的吸收率为68.93%。
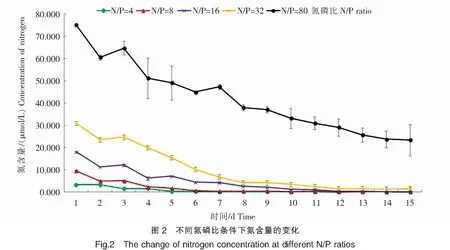
2.3 不同氮磷比条件下米氏凯伦藻对磷的消耗特征
米氏凯伦藻对磷的吸收特征如图3所示,前3天各组培养液中的磷浓度均持续下降,而后由于受氮浓度的影响,第4天开始各组米氏凯伦藻对磷的吸收特征有所不同。在N/P=4条件下,磷的消耗主要发生在前3天,第4天开始磷含量就趋于平稳,且保持较高水平,在0.600 0 μmol/L附近浮动,至第15天实验结束时,培养液中磷含量减少了0.229 0 μmol/L,吸收率为28.17%;当N/P=8时,前5天磷含量快速减少,降低至0.387 1 μmol/L,之后磷水平一直维持在该水平,但是在第8天时磷含量又开始上升,上升至约0.600 0 μmol/L水平,并维持在该水平,至实验结束时,培养液中磷含量降低了0.256 5 μmol/L,吸收率为31.86%;在N/P=16条件下,磷的消耗主要是在前5天,在第6天时磷含量降低至0.196 8 μmol/L,并一直维持在该水平,至实验结束时,磷含量减少了0.532 3 μmol/L,吸收率为69.18%;在N/P=32条件下,前6天磷含量快速下降,在第7天时下降至0.227 4 μmol/L,此后一直维持在该水平,至第15天时磷含量减少了0.622 6 μmol/L,吸收率为77.05%;当N/P=80时,前10天磷含量大量下降,在第10天磷含量降至0.209 7 μmol/L,之后基本维持在该水平,至第15天实验结束时,培养液中磷酸盐含量减少了0.637 1 μmol/L,磷吸收率为79.16%。
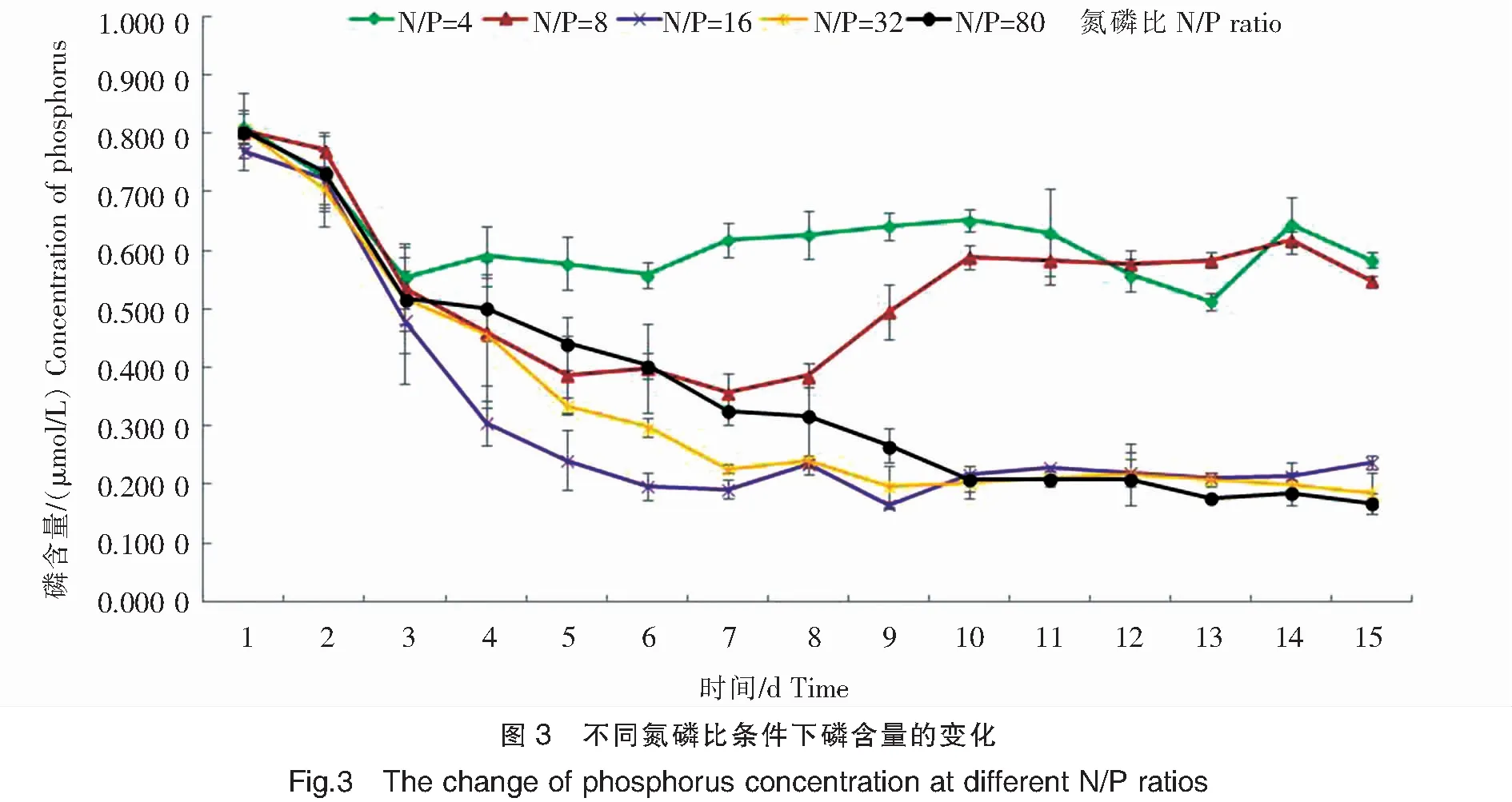
2.4 米氏凯伦藻生长过程中氮磷比的变化
从图4中可以看出,除了N/P=80组外,其他各组的氮磷比的总体变化趋势为逐渐下降,具体可分为两个阶段,第一个阶段是在停滞期和对数生长期,由于氮和磷同时被消耗,氮磷比下降幅度小,甚至个别组还出现高于初始氮磷比的现象;但是到了第6~7天,此时各组细胞均已接近最高细胞数,但培养液中的氮磷比均开始逐渐下降,直至实验结束时,各组氮磷比均在10以下,N/P为4、8和16组甚至接近0。而在N/P=80条件下,由于磷限制问题,藻类对氮的吸收有限,导致氮磷比未有明显下降,反而有所升高。
同时从闽东监测中心提供的2012年三沙湾米氏凯伦藻赤潮发生海域监测的营养盐数据来看,当三沙湾海域米氏凯伦藻密度超过赤潮基准浓度(103个/mL)形成赤潮时,发生海域的氮磷比值较低,变化范围为0.14~33.86,绝大部分监测点氮磷比比值在16以下。

3 讨论
氮和磷是浮游植物生长所需的必要元素,营养盐结构的改变将会导致浮游植物群落组成的改变以及赤潮的频发[5-6]。但是不同的藻类对于营养盐的需求不同,因此每一种浮游藻类的最适氮磷比也有不同。本次实验结果表明除了N/P=4外,米氏凯伦藻在其它氮磷浓度比条件下均生长良好,与对照组相比存在极显著差异。在中低氮磷比(N/P为4、8、16)条件下,米氏凯伦藻能够较快进入对数期,并且达到最高细胞数,但之后细胞数随即下降,提前进入衰亡期;通过检测培养液中氮和磷的浓度可知,在中低氮浓度组的对数期,米氏凯伦藻消耗了大量的氮,氮的吸收率均接近100%,而磷浓度的消耗主要在停滞期,进入对数期后磷浓度则变化不大。可见,在中低氮磷比条件下,米氏凯伦藻前期迅速耗尽了营养盐,较早地进入对数期并且达到最高细胞数,但是对数期后较低含量的氮浓度无法继续支持细胞的继续生长和繁殖,所以藻细胞数迅速下降,提前进入衰亡期。而在高氮磷比(N/P为32、80)条件下,米氏凯伦藻推迟进入对数期,可能是由于前期过高的氮浓度单盐毒害,抑制了藻细胞的分裂,而经过数天的抑制期,氮浓度相对降低后,藻类才得以进入对数期[7];但在该条件下,藻类达到最高细胞数后并没有直接进入衰亡期,而是进入平稳期,维持着较高的细胞密度。通过检测培养液中氮和磷的浓度可知,在高氮磷比条件下,米氏凯伦藻在对数生长期主要消耗的是磷,而对于氮的消耗远低于中低氮磷比组。相关研究表明米氏凯伦藻对氮的需求高,不仅在对数生长期对氮的吸收利用较高,在培养后期仍然会持续利用[8]。本次实验结果也显示,米氏凯伦藻在氮磷比值较低的情况下,虽然前期消耗大量的氮浓度以快速进入对数期,但却由于后期氮含量耗尽提前进入衰亡期;在高氮磷比值条件下,虽然前期磷被大量消耗,但中后期氮含量仍然较高,藻类能够维持较长时间的高密度状态,可见米氏凯伦藻对于氮的需求高于磷,适当的高氮环境更适合米氏凯伦藻生长。
Klausmeier等利用模型得出浮游植物的最适氮磷比是根据生态条件的变化而变化,变化范围在8.2~45.0之间,还指出Redfield系数(N/P=16)并非普遍的生化最适氮磷比,而是代表了平均水平[9];曹春晖等[7]的研究结果表明米氏凯伦藻在氮磷比为15∶1到276∶1之间均可达到最高细胞密度和相对生长率;孙军等[10]指出米氏凯伦藻在氮磷比为80的条件下比生长率最大;斯丹等[11]的研究结果认为米氏凯伦藻的最适氮磷比为50;而马龙等[12]认为较高或较低的N/P均不适合米氏凯伦藻生长,该藻的最适氮磷比应该为24.4。本次实验结果可知,米氏凯伦藻在氮磷比为8到32之间,其最高细胞数和最大比生长率随着氮磷比的增加而增加,当氮磷比为80时,虽然最高细胞数高于其他组,但是最大比生长率仅为0.20 d-1;过高或过低的氮磷比均不适合米氏凯伦藻的生长,其最适氮磷比为32,与马龙等[12]实验结果较为接近。Redfield定律认为,如果氮磷比高于16∶1,则考虑磷为限制因素,而当氮磷比小于10∶1时,则认为氮为限制因素;当氮磷比比值在10~20之间时,限制性因素则变得不确定[13]。综合本次实验结果和其他研究成果可见,米氏凯伦藻最适氮磷比均高于16∶1,更适合生长于磷限制潜在的生态环境中。
同时,将本次实验结果与2012年三沙湾米氏凯伦藻赤潮现场数据相比较发现,现场氮磷比比值均较低,最小值仅为0.14。龙华等[14]在2003年连江米氏凯伦藻赤潮现场监测的氮磷比比值也偏低,多数小于16,最低值仅为0.4,认为低氮磷比是米氏凯伦藻赤潮形成的重要原因,与2012年三沙湾现场调查结果接近。但现场调查往往是在赤潮发生、海水开始变色后才开展的,此时米氏凯伦藻的密度均在103个/mL以上,已逐渐接近最高密度,而根据实验室结果发现,当藻类细胞数接近最高密度时,培养液中的氮磷比比值已经开始下降,至赤潮结束时,氮磷比比值已大幅下降,有的甚至接近于0。因此本文认为,造成实验室米氏凯伦藻生长最适氮磷比与赤潮暴发现场氮磷比的差异巨大的原因在于,赤潮现场测定的氮磷浓度是在米氏凯伦藻大量吸收利用营养盐后测定的,周围环境中的硝酸盐含量的消耗大于磷酸盐,从而导致了氮磷比下降。该现象在姚炜民等[15]在浙江米氏凯伦藻赤潮海域的连续监测结果中也有出现:在赤潮未发生时,N/P比值为24.2,赤潮维持期间N/P比值为12.5,赤潮消亡期间N/P比值为12.8,赤潮期间氮磷比显著低于初始氮磷比。综上所述,低氮磷比并非是米氏凯伦藻形成的重要原因,而是赤潮发生期间,米氏凯伦藻消耗了大量的营养盐,氮含量大幅下降造成的结果。
4 结论
1)分离自福建沿海的米氏凯伦藻在氮磷比比值为4~80之间均能生长,氮磷比对米氏凯伦藻的生长有显著影响,过高或过低的氮磷比均不适合米氏凯伦藻生长,其最适生长氮磷比比值为32。在该条件下,米氏凯伦藻的最高细胞数为2.68×103个/mL,最大比生长率为0.33 d-1。
2)米氏凯伦藻对氮的需求高于磷,整个生长周期都会持续利用吸收氮,在适当的磷限制环境中,米氏凯伦藻能够维持较长的生长周期。
3)与现场测定的营养盐数据相比较,发现低氮磷比并不是米氏凯伦藻赤潮形成的原因,而是赤潮发生期间,米氏凯伦藻消耗大量营养盐,氮含量大幅下降的结果。
[1]国家海洋局.2012年中国海洋灾害公报[EB/OL].[2013-03-11].http://www.coi.gov.cn/gongbao/zaihai/201303/t20130311_26237.html
[2]黄凯旋.米氏凯伦藻的氮营养生理生态研究[D].广州:暨南大学,2007.
[3]李铁,胡立阁,史致丽.营养盐对中肋骨条藻和新月菱形藻生长及氮磷组成的影响[J].海洋与湖沼,2000,31(1):46-52.
[4]Stéphane L H,Christian M,Pierre L C.Nitrogen uptake in permanently well-mixed temperate coastal waters[J].Estuarine,Coastal and Shelf Science,1996,42(6):803-818.
[5]Hodgkiss I J,Ho K C.Are changes in N∶P ratios in coastal waters the key to increased red tide blooms?[J].Hydrobiologia,1997,352(1-2):141-147.
[6]Li J,Glibert P M,Alexander J A.Effect of ambient DIN∶DIP ratio on the nitrogen uptake of harmful dinoflagellateProrocentrumminimumandProrocentrumdonghaiensein turbidistat[J].Chinese Journal of Oceanology and Limnology,2011,29(4):746-761.
[7]曹春晖,刘文岭,施定基,等.不同氮磷浓度对米氏凯伦藻生长的影响[J].天津科技大学学报,2010,25(2):22-25.
[8]沈盎绿,李道季.不同营养盐水平对东海原甲藻和米氏凯伦藻生长的影响[J].海洋渔业,2016,38(4):415-422.
[9]Klausmeier C A,Litchman E,Daufresne T,et al.Optimal nitrogen-to-phosphorus stoichiometry of phytoplankton[J].Nature,2004,429(6988):171-174.
[10]孙军,刘东艳,陈宗涛,等.不同氮磷比率对青岛大扁藻、新月柱鞘藻和米氏凯伦藻生长影响及其生存策略研究[J].应用生态学报,2004,15(11):2122-2126.
[11]斯丹,张拿慧,刘桂群,等.不同氮磷比对米氏凯伦藻和中肋骨条藻种群竞争的影响[J].海洋湖沼通报,2016,(5):118-124.
[12]马龙,王培刚,朱葆华,等.米氏凯伦藻与亚心形扁藻在不同营养盐水平下的种间竞争[J].海洋环境科学,2013,32(2):221-226.
[13]丰茂武,吴云海,冯仕训,等.不同氮磷比对藻类生长的影响[J].生态环境,2008,17(5):1759-1763.
[14]龙华,杜琦.福建沿海米氏凯伦藻赤潮的初步研究[J].福建水产,2005,12(4):22-26.
[15]姚炜民,潘晓东,华丹丹.浙江海域米氏凯伦藻赤潮成因的初步研究[J].水利渔业,2007,27(6):57-76.
The effects of nitrogrn to phosphorus ratio on the growth ofKareniamikimotoiseparated from Fujian coastal area
PAN Feifei,WU Nan
(Monitoring Center of Marine Environment and Fishery Resources,Fuzhou 350003,China )
The growth character ofKareniamikimotoiwas studied in culture medium at different ambient ratios of N/P(4∶1,8∶1,16∶1,32∶1,80∶1).The results showed that N/P ratio significantly affected the growth ofKareniamikimotoi.Too high or too low N/P ratio was not suitable forKareniamikimotoigrowth.The algae grew fastest under 32 N/P ratios,and the maximum specific growth rate was 0.33 d-1.Kareniamikimotoidemanded more nitrogen than phosphorus for growth,and it could maintain a longer growth cycle in the appropriate phosphorus limited environment.The main reason ofKareniamikimotoired tides was not the low N/P ratio in the environment,but was a large number of nutrient consumption,especially nitrogen.
N/P ratio;red tide;Kareniamikimotoi
2017-05-12
福建省海洋与渔业结构调整专项资金(闽财指〔2016〕841号文).
潘非斐(1987-),女,工程师,硕士,研究方向:海洋环境.Tel:18695602045.E-mail: 782494772@qq.com
P735
A
1006-5601(2017)04-0287-08
潘非斐, 吴 楠.不同氮磷比对福建沿海米氏凯伦藻生长的影响[J].渔业研究,2017,39(4):287-294.
