中肋骨条藻-裸甲藻-双毛纺锤水蚤营养传递的级联效应
2020-06-03张翠霞李世玥吴宇佳吴梦瑶刘泽洋刘宇轩杨昊达
张翠霞 , 高 歌 李世玥 吴宇佳 王 蕊 吴梦瑶 刘泽洋 刘宇轩杨昊达
(1. 天津科技大学 海洋与环境学院, 天津 300457; 2. 天津市海洋资源与化学重点实验室, 天津 300457; 3. 天津市海洋环境保护与修复技术工程中心, 天津 300457)
传统的观点认为, 浮游生物食物链就是从自养的浮游植物利用无机的营养盐, 被浮游动物摄食,然后再传递到更高的营养层次。后来随着微食物环的发展, 微型浮游动物即原生动物被考虑进来, 成为联系浮游植物和浮游动物的纽带[1]; 微型浮游动物对浮游植物有很强的摄食压力[2], 尤其是在藻华期间[3-4]。如果高营养级的中型浮游动物如桡足类因摄食了微型浮游动物, 从而释放微型浮游动物对浮游植物的摄食压力, 使得浮游植物增加, 被称为营养级联(TC, trophic cascade)效应。相比较于来自中型浮游动物直接的摄食死亡, 间接的营养级联效应增加浮游植物丰度, 降低中型浮游动物的摄食率, 从而弱化浮游生物网营养传递过程中的下行控制(topdown)的作用[5-6]。
甲藻是微型浮游动物的重要组成部分, 营混合营养方式, 通过多种策略摄食不同类型和不同粒级大小的生物, 在浮游生物网的营养传递和生物泵过程中发挥着多重角色的作用[7-9]。甲藻对食物的摄食选择不仅跟种类有关, 还跟大小有关, 摄食者与食物的大小比例大约是0.4∶1到7∶1之间[10]。裸甲藻(Gymnodinium sp.)和中肋骨条藻(Skeletonema costatum)都是我国黄渤海浮游植物典型的代表物种[11-12], 经常是形成赤潮的原因物种[13-14]。裸甲藻能够通过吞噬营养的方式来选择摄食隐藻、甲藻和等鞭金藻等[15-16]。Yoo等[16]用透射电镜证实了裸甲藻能够摄食中肋骨条藻, 并通过室内实验研究裸甲藻对中肋骨条藻的摄食率。
营养级联效应越来越引起生态学家的重视, 已有的研究大多用来定性分析或解释摄食问题和实验现象[17-19], 但是营养级联效应的大小和对浮游植物的具体影响还不是很清楚, 这方面的研究还相对较少[6,20]。因此, 对营养级联效应的研究便于更深入认识复杂的食物营养传递过程。本研究选择渤海湾桡足类的典型代表物种双毛纺锤水蚤(Acartia bifilosa)作为中型浮游动物, 以裸甲藻和中肋骨条藻分别作为微型浮游动物和浮游植物。通过实验室内培养模拟食物链中肋骨条藻-裸甲藻和食物链中肋骨条藻-裸甲藻-双毛纺锤水蚤, 来定量地分析桡足类的摄食选择性和营养级联效应的大小以及对浮游动物摄食浮游植物的影响, 为自然水体藻华和赤潮的发生和控制提供重要的科学依据和基础。
裸甲藻的等效球体直径(ESD, equivalent spherical diameter)是 15 μm, 中肋骨条藻的 ESD 为 5.3 μm,两者单细胞的大小比例是2.8。为避免实验过程中裸甲藻的自养方式对摄食中肋骨条藻的影响, 实验前将中肋骨条藻藻液作为食物与裸甲藻混合, 加入仅能维持中肋骨条藻生长的f/2培养基, 进行驯化培养。
1 材料与方法
1.1 实验准备
1.1.1 藻种的培养与保存
于天津近岸海域(北塘海域)取海水, 使用中性滤纸和普通微孔滤膜(0.8 μm)依次过滤海水, 并将其高压灭菌, 配置f/2培养基。在光照培养箱中, 设置适宜的条件: 光照强度2 000 lx, 光照周期12∶12, 温度18, ℃培养实验所需要的纯种中肋骨条藻和裸甲藻的藻液。
1.1.2 桡足类的采集
于天津近岸海域(北塘海域)使用型号为 PTNS120的浮游生物网(孔径 112 μm)拖网, 收集桡足类保存于500 mL~1 L的玻璃培养瓶中; 立刻带回到实验室, 转移到培养槽中, 添加裸甲藻和中肋骨条藻,置于光照培养箱里, 设定温度18, ℃暗光条件下曝气进行驯化培养。
1.1.3 裸甲藻和中肋骨条藻混合藻液的配置和驯化培养
实验前将纯种的藻液混合配置两份450 mL的混合藻液, 中肋骨条藻的密度 8 273 cells/mL, 裸甲藻密度2 430 cells/mL, 只加入100 mL f/2培养基维持中肋骨条藻的生长。实验前两天停止向混合藻液中加入中肋骨条藻, 将上层清液转移出得到高密度的裸甲藻和低密度的中肋骨条藻的混合藻液。
1.1.4 细胞生物量
通过细胞平均体积估算中肋骨条藻和裸甲藻的碳含量[21], 中肋骨条藻的碳生物量是0.010 ng C/cell,裸甲藻的碳生物量是0.347 ng C/cell。
1.2 实验的设计(表1)
1.2.1 裸甲藻摄食中肋骨条藻
在中肋骨条藻处于指数生长期时, 分别设置 6组不同的中肋骨条藻密度梯度(处理组编号为 1—6,表1), 每个处理组设置3个平行样, 裸甲藻的密度设置为100 cells/mL(生物量34.7 ng C/mL), 每个对照组设置两个平行样(对照组不添加裸甲藻)。藻液加入到 125 mL的聚碳酸酯培养瓶中, 最后添加浓度为f/20的培养基维持中肋骨条藻的生长。所有的培养瓶置于光照培养箱中培养, 每6小时摇匀一次, 于48 h后进行裸甲藻和中肋骨条藻的取样。

表1 中肋骨条藻、裸甲藻和双毛纺锤水蚤摄食培养实验的初始密度及生物量设置Tab. 1 Initial concentration and biomass of Skeletonema costatum, Gymnodinium sp., and Acartia bifilosa in feeding experiments
1.2.2 桡足类摄食裸甲藻和中肋骨条藻
同时使用 500 mL的 Nalgene聚碳酸酯培养瓶,设置5个不同的中肋骨条藻密度梯度(处理组编号为A—E, 表 1), 每个处理组设置 3个平行样。裸甲藻的密度设置为 100 cells/mL(生物量 34.7 ng C/mL),挑取12只健康活泼的桡足类优势物种双毛纺锤水蚤添加到每个培养瓶中。渤海湾天津近岸海域记录的双毛纺锤水蚤最大丰度是34 ind./L[22], 本实验设置双毛纺锤水蚤的初始丰度为 24 ind./L。设置不添加桡足类的5个对照组, 每个对照组2个平行。每个培养瓶加入50 mL的f/2培养基支持中肋骨条藻的生长。所有的培养瓶置于光照培养箱中培养, 每6小时摇匀一次, 于48 h后观察并记录桡足类的生存状况,进行裸甲藻和中肋骨条藻取样。
1.2.3 实验取样和样品分析
两组实验同时进行培养, 48 h后, 每个培养瓶取样品 10 mL置于离心管中, 用鲁格试剂固定避光阴凉处保存。取1 mL样品, 用光学显微镜观察中肋骨条藻细胞并计数; 取3 mL样品, 用倒置显微镜观察裸甲藻的细胞并计数。
1.3 数据处理与分析

P0和 Pt为培养前后裸甲藻的密度, t为培养的时间;
不同处理组裸甲藻的生长率(μ)拟合米氏方程:
1.3.1 裸甲藻摄食中肋骨条藻的实验
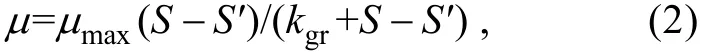
式中, μmax为最大生长率(单位: d-1), S为培养后单位体积的中肋骨条藻的生物量(单位: ng C/mL), S′为中肋骨条藻的初始生物量(单位: ng C/mL), Kgr为米氏常数,当 μ=1/2μmax时的中肋骨条藻的生物量(单位: ng C/mL)。
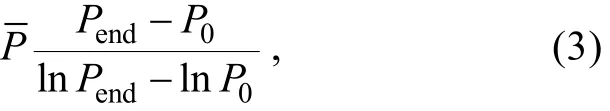
清滤率(F, filterance/clearance rate)[23]:

g: 摄食率(单位: d-1);
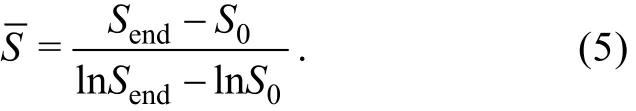

裸甲藻的摄食率(I, ng C/d)拟合米氏(Michaelis-Menten)方程:

Imax: 最大摄食率(单位: ng C/d ), Kir: 当 I=1/2 Imax时的中肋骨条藻的生物量(单位: ng C/mL);
将公式(7)代入到公式(6)得到裸甲藻的摄食率(g,单位: d-1)的公式(8), 与裸甲藻和中肋骨条藻的培养过程中的平均生物量有关。

1.3.2 添加桡足类的摄食实验
桡足类对裸甲藻的摄食率(grazing rate, Gc, 单位: d-1)[24]:

pt: 处理组裸甲藻的终生物量; pc: 对照组裸甲藻的终生物量; t: 培养时间;
清滤率(clearance rate, FC, 单位: mL/d)[24]:

V(mL): 培养瓶的水体体积; n: 处理组添加桡足类的数量;
摄食率(ingestion rate, IC, 单位: ng C/d)[24]:

P: 裸甲藻的平均生物量(单位: ng C/mL);
对照组中肋骨条藻的净生长率(control, kc, 单位:d-1):

g(单位: d-1): 对照组裸甲藻对中肋骨条藻的摄食率, 由公式(8)计算可得;
处理组中肋骨条藻的净生长率(treatment, kt, 单位: d-1:

g′(d-1): 处理组桡足类存在时, 裸甲藻对中肋骨条藻的摄食率, 由公式(8)计算可得;
公式(12)代入公式(13)得到桡足类对中肋骨条藻的直接摄食(死亡)率(grazing rate, Ga, 单位: d-1):
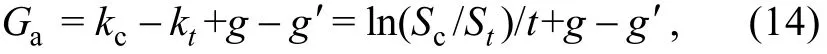
St(单位: ng C/mL): 处理组中肋骨条藻的终生物量; Sc(单位: ng C/mL): 对照组中肋骨条藻的终生物量。
营养级联效应(TC, the rate of the trophic cascade)[20]:

摄食选择指数(Feeding selectivity index, α)为摄食碳量占总的饵料碳量的百分比[25]。该指数介于0~1,当 α>0.5时, 表明具有摄食的偏好, 当 α>0.5时, 表明没有摄食的偏好。通过Origin 9.0拟合裸甲藻的生长率、摄食率与中肋骨条藻生物量的Michaelis-Menten方程曲线, 统计分析通过IBM SPSS Statistics 19.0实现。
2 结果
2.1 裸甲藻的生长率和摄食率
在裸甲藻摄食中肋骨条藻的实验中, 裸甲藻的生长率随着中肋骨条藻生物量变化的增加而增大(图1), 在中肋骨条藻生物量为250~750 ng C/mL增长速率最大, 而后趋于平缓。通过拟合 Michaelis-Menten方 程 μ =0.52(S -S ′ )/(146.74+ S - S ′), 裸甲藻的最大生长率(μmax)为0.52 d-1, 米氏常数kgr显示, 其半饱和细胞生物量为146.7 ng C/mL。

图1 裸甲藻的生长率(μ, d-1)与中肋骨条藻的生物量变化(ng C/mL)之间的关系, 曲线代表拟合的米氏方程曲线, 误差棒分别代表生长率和生物量变化的标准偏差Fig. 1 Specific growth rate (μ, d-1) of Gymnodinium sp.feeding on Skeletonema costatum as a function of change in prey biomass (ng C/mL); the curve represents the fitting curve of the Michaelis-Menten equation, whereas the error bars represent the standard deviations of growth rate and biomass change
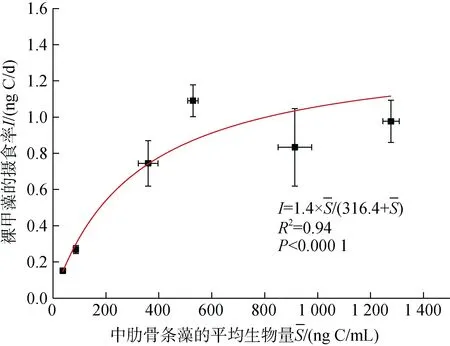
图 2 裸甲藻的摄食率(ng C/d)与中肋骨条藻平均生物量(ng C/mL)的关系, 曲线代表拟合的米氏方程曲线,误差棒分别代表摄食率和平均生物量的标准偏差Fig. 2 Ingestion rates (ng C/d) of Gymnodinium sp. on Skeletonema costatum as a function of prey mean concentration (ng C/mL); the curve represents the fitting curve of the Michaelis-Menten equation, whereas the error bars represent the standard deviations of ingestion rate and mean concentration
裸甲藻的摄食率与中肋骨条藻的平均细胞生物量成显著正相关(P < 0.000 1), 中肋骨条藻低生物量条件下摄食率增加较快, 高生物量的情况下增加较缓慢(图2)。通过拟合 Michaelis-Menten方程得到裸甲藻对中肋骨条藻的最大摄食率(Imax)为1.4 ng C/d, 半饱和生物量为316.4 ng C/mL。从而得到裸甲藻摄食率的计算公式为后面添加桡足类培养实验研究营养级联效应提供了基础。
2.2 双毛纺锤水蚤的清滤率和摄食率
当中肋骨条藻的平均生物量为93.19 ng C/mL时,双毛纺锤水蚤对裸甲藻的清滤率为最大41.53 mL/d,同时对中肋骨条藻的清滤率也为最大 26.90 mL/d;在中肋骨条藻高生物量时, 双毛纺锤水蚤对裸甲藻和中肋骨条藻的清滤率都呈现下降(图3)。双毛纺锤水蚤对裸甲藻的清滤率显著高于对中肋骨条藻的清滤率(成对样本 t检验, P<0.001, df=4)。
随着中肋骨条藻生物量的增加, 双毛纺锤水蚤对中肋骨条藻的摄食率(479.22~4 511.61 ng C/d)逐渐增加,在中肋骨条藻的平均生物量最高为707.04 ng C/mL (处理组E)时, 摄食率达到最大值。双毛纺锤水蚤对裸甲藻的摄食率介于682.78 ~ 772.00 ng C/d, 呈先增加后降低的趋势; 在中肋骨条藻的平均生物量为93.19 ng C/mL(处理组 C)时, 摄食率最大值为772.00 ng C/d(图 4)。
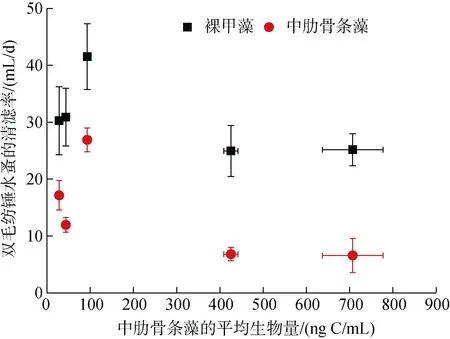
图 3 不同食物生物量下双毛纺锤水蚤对裸甲藻(方形)和中肋骨条藻(圆形)的清滤率, 误差棒分别代表清滤率和平均生物量的标准偏差Fig. 3 Clearance rate (mL/d) of Acartia bifilosa on Gymnodinium sp (square) and Skeletonema costatum(circle) in different mean concentrations of Skeletonema costatum; the error bars represent the standard deviations of the clearance rate and mean concentration
2.3 双毛纺锤水蚤的摄食选择性
低生物量的A和B两个处理组, 双毛纺锤水蚤对裸甲藻的摄食选择指数大于对中肋骨条藻的选择指数, 且数值都大于 0.5, 表明对裸甲藻具有摄食的偏好性; 而在高生物量的处理组(C、D和E), 双毛纺锤水蚤对中肋骨条藻的摄食选择指数(0.76~0.85)都大于 0.5, 对裸甲藻的摄食选择指数偏低, 表明对中肋骨条藻具有摄食的偏好性(图5)。

图4 不同处理组双毛纺锤水蚤对裸甲藻(黑色)和中肋骨条藻(红色)的摄食率, 误差棒代表其标准偏差Fig. 4 Ingestion rate (in ng C/d) of Acartia bifilosa in different treatments. Black bars are ingestion rates for Gymnodinium sp, red bars are ingestion rates for Skeletonema costatum; the error bar represents the standard deviation
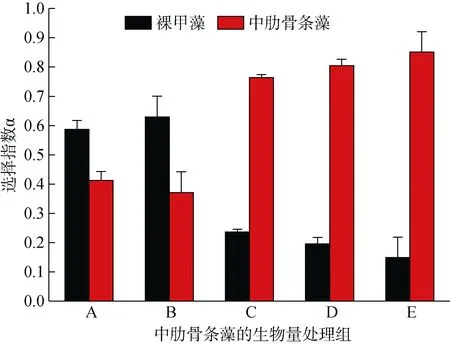
图 5 不同处理组双毛纺锤水蚤对裸甲藻(黑色)和中肋骨条藻(红色)的摄食选择指数, 误差棒代表其标准偏差Fig. 5 Feeding selectivity index (α) of Acartia bifilosa in different treatments; black bars indicate the indexes for Gymnodinium sp, whereas red bars are indexes for Skeletonema costatum; the error bar represents the standard deviation
2.4 营养级联效应
在食物链双毛纺锤水蚤-裸甲藻-中肋骨条藻的营养传递过程中, 在不同的中肋骨条藻生物量下营养级联效应(TC)都为正值, 介于 0.018~0.12 d-1, 随中肋骨条藻细胞生物量的增加, 营养级联效应呈现降低, 在高生物量组的变化较小(图 6)。双毛纺锤水蚤对中肋骨条藻的直接摄食死亡率(Ga)用负值来表示, 范围介于-0.75~-0.10 d-1, 绝对值都高于营养级联效应, 并且中肋骨条藻低生物量组的直接摄食死亡率绝对值高于高生物量的处理组。
In(St/Sc)/t代表添加双毛纺锤水蚤后, 中肋骨条藻细胞生物量对桡足类的对数响应, 范围介于-0.70~-0.07 d-1, 是营养级联正效应和双毛纺锤水蚤对中肋骨条藻的直接摄食死亡负效应两者的综合结果。添加双毛纺锤水蚤后, 直接摄食死亡明显大于营养级联效应, 导致相比较不添加双毛纺锤水蚤时中肋骨条藻生物量没有增加反而降低。中肋骨条藻低生物量处理组的对数变化值的绝对值最大, 在高生物量时, 绝对值减小接近0。中肋骨条藻的直接摄食死亡率和其细胞生物量的对数响应, 都为负值, 随中肋骨条藻的生物量的变化趋势是相似的(图 6), 因此双毛纺锤水蚤对中肋骨条藻的直接摄食主要决定了中肋骨条藻的最终生物量变化。

图 6 双毛纺锤水蚤对中肋骨条藻的直接摄食(死亡)率(Ga)、营养级联效应(TC)和中肋骨条藻细胞生物量对桡足类的对数响应[In(St/Sc)/t]与中肋骨条藻平均生物量(ng C/mL)的关系Fig. 6 Direct grazing mortality (Ga) on Skeletonema costatum by Acartia bifilosa, rate of trophic cascades(TC) and [In(St/Sc)/t] of Skeletonema costatum, as a function of mean diatom concentration (ng C/mL)
3 讨论
3.1 桡足类的摄食选择性
在裸甲藻摄食中肋骨条藻的实验中, 不考虑两者之间的化感作用, 裸甲藻的摄食率随中肋骨条藻平均生物量的变化与 Michaelis-Menten方程的拟合度较高(图 2, R2=0.94, P<0.000 1), 满足添加桡足类摄食实验的要求。
双毛纺锤水蚤对中肋骨条藻的摄食选择系数随其平均生物量呈现显著指数增长(R2=0.8, P<0.000 1),而裸甲藻平均生物量对双毛纺锤水蚤的摄食选择系数影响不明显(图7)。中肋骨条藻的生物量是决定双毛纺锤水蚤摄食选择性的重要因素, 并且生物量越高, 双毛纺锤水蚤对中肋骨条藻的选择性越强。
对照天津市近岸实际海域的浮游植物丰度来看,本实验设置的两个低生物量处理组的中肋骨条藻细胞丰度都大于冬季的最低值[26], 应该满足浮游动物的基本摄食需求。B处理组的双毛纺锤水蚤对裸甲藻和中肋骨条藻的摄食选择差异要大于 A处理组, 可以看出双毛纺锤水蚤的摄食是主动选择的行为。从食物质量来看, 裸甲藻能为双毛纺锤水蚤提供大量的生长、繁殖和发育必需的二十二碳六烯酸(docosahexaenoic acid, DHA), 相比较中肋骨条藻有更多的营养价值[27]。大量的室内和现场实验研究证实, 当硅藻和甲藻同时存在时, 桡足类更倾向于摄食甲藻[28-31]。在中肋骨条藻低生物量的条件下, 即食物生物量不充足的条件下, 裸甲藻能为双毛纺锤水蚤提供更多的营养来源, 这是双毛纺锤水蚤选择摄食裸甲藻的原因。

图7 双毛纺锤水蚤的摄食选择指数与中肋骨条藻(a)和裸甲藻(b)的平均生物量关系Fig. 7 Change in feeding selectivity index of Acartia bifilosa with prey Skeletonema costatum (a) and Gymnodinium sp. (b)mean biomass
在高生物量的处理组, 即食物生物量充足或饱和状态下, 双毛纺锤水蚤选择性摄食中肋骨条藻,对其摄食率显著高于对裸甲藻摄食率; 在最高生物量的处理组, 双毛纺锤水蚤对中肋骨条藻摄食率比对裸甲藻的摄食率高出6倍以上(图4)。该实验的结果与之前的研究[20]不同, 可能跟桡足类和甲藻的物种不同有关。纺锤水蚤属是近岸常见的桡足类物种,偏向于摄食大的颗粒物质[32-33]。实验过程中肋骨条藻多数是以链状形式的存在, 每条链有 10~30个细胞不等, 不同长度的细胞链对于桡足类在颗粒大小的选择上优于裸甲藻, 同时裸甲藻还可以通过自身的运动能力躲避双毛纺锤水蚤的猎食。食物组成结构也可能是影响摄食选择性的一个重要因素, 本研究中中肋骨条藻和裸甲藻初始生物量跟已有的研究[20]不同。裸甲藻的细胞偏大, 单细胞的碳含量高于其他甲藻, 整体上裸甲藻和中肋骨条藻初始碳含量的比值较高。桡足类的摄食选择性会随着微型浮游动物的生物量而变化[6], 因此食物碳含量组成对选择性摄食的影响还需要后续进一步的实验来研究。此外, 裸甲藻作为代表性的赤潮原因物种, 能够分泌一种神经性贝毒(Neurotoxic shellfish poisoning, NSP)的毒素, 会对捕食者具有一定的毒性。当有不同种类的可供选择的混合食物时, 桡足类会主动避开摄食有毒的甲藻, 会选择摄食无毒的硅藻[34]。在不同生物量下双毛纺锤水蚤摄食行为的这种差异, 虽然不同于已有的部分研究[20,29],但是符合许多现场实验支持的一般模式, 这种模式认为在藻华末期或水体分层期的浮游植物生物量较低的情况下, 中型浮游动物一般偏向于摄食微型浮游动物[35-36], 而在藻华期浮游植物生物量较高时, 中型浮游动物更偏向于摄食大小适合的植物细胞[18,37-38]。本研究过程观察到中肋骨条藻呈现不同长度的细胞链, 可为双毛纺锤水蚤营造出不同大小的食物颗粒。
3.2 营养级联效应及对浮游植物的影响
本研究的营养级联效应都为正值, 说明添加的双毛纺锤水蚤摄食了裸甲藻, 减少了裸甲藻对中肋骨条藻的摄食, 从而间接地降低了双毛纺锤水蚤和裸甲藻对中肋骨条藻的摄食控制作用。营养级联效应与裸甲藻对中肋骨条藻的摄食率呈显著的正相关(图 8, 双侧Pearson相关性分析, R2=0.74, P<0.01), 低生物量下的裸甲藻对中肋骨条藻的摄食率大于高生物量的处理组, 裸甲藻的摄食率随生物量的升高而降低, 因此营养级联效应也呈现降低的趋势(图6)。

图 8 营养级联效应(TC, 单位: d-1)与裸甲藻对中肋骨条藻摄食率(g′, 单位: d-1)的相关关系Fig. 8 Correlation between the rate of trophic cascades and ingestion rate on the grazing rates of Gymnodinium sp. by Skeletonema costatum
研究发现营养级联效应与双毛纺锤水蚤对裸甲藻和中肋骨条藻的摄食选择指数呈现极显著的线性相关(图9, R2=0.85, P<0.001), 这说明营养级联效应与双毛纺锤水蚤的摄食行为密切相关。营养级联效应随着双毛纺锤水蚤对裸甲藻的摄食选择指数的变大而增大, 在中肋骨条藻低生物量的情况下, 双毛纺锤水蚤对裸甲藻的摄食率略大于对中肋骨条藻的摄食率, 但是双毛纺锤水蚤更偏好于选择摄食裸甲藻(图 4), 从而降低了对中肋骨条藻的摄食压力, 表现出高的营养级联效应。然而在中肋骨条藻高生物量的情况下, 双毛纺锤水蚤偏向于直接选择摄食中肋骨条藻, 降低了对裸甲藻的摄食率, 原因可能来自于中肋骨条藻高的细胞密度降低了双毛纺锤水蚤的清滤率(图3), 由于裸甲藻的细胞生物量相对较低,因此高细胞生物量对裸甲藻起到了庇护的作用[39],导致营养级联效应降低。
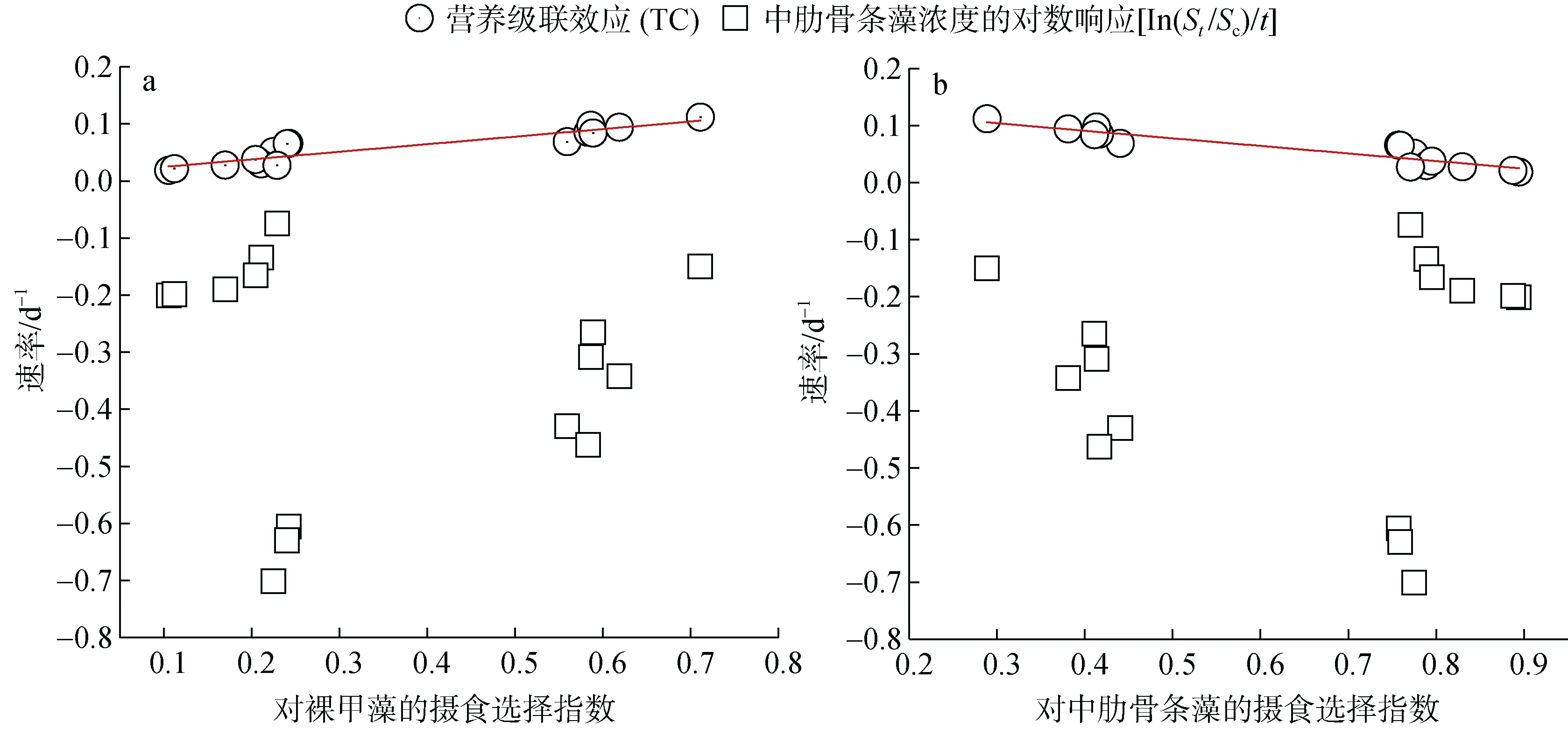
图9 不同的速率与双毛纺锤水蚤对裸甲藻(a)和中肋骨条藻(b)的摄食选择指数的相关关系, 营养级联效应与摄食选择指数拟合线性回归, R2=0.85, P<0.001Fig. 9 Relationship between the different rates of Skeletonema costatum and feeding selectivity indexes on Gymnodinium sp.(a) and Skeletonema costatum (b). Linear regression: R2 = 0.85, P<0.001
培养前后中肋骨条藻的对数响应[In(St/Sc)/t]取决于双毛纺锤水蚤的直接摄食(负效应)和营养级联效应(正效应)两者的平衡。本研究中的[In(St/Sc)/t]与双毛纺锤水蚤的摄食选择指数没有显著的相关性(图9), 由于第三个处理组的点比较离散, 若把第三个处理组的点去掉, 两者呈现弱的线性相关(R2=0.28,P<0.01)。在中肋骨条藻低生物量的情况下, 双毛纺锤水蚤既选择摄食裸甲藻, 也直接摄食中肋骨条藻,但是对中肋骨条藻的直接摄食死亡率显著高于营养级联效应; 高的中肋骨条藻生物量下, 直接摄食死亡率的绝对值与营养级联效应相反, [In(St/Sc)/t]接近于0。双毛纺锤水蚤对中肋骨条藻的摄食率随着其生物量的增加而显著增加(图5, 单样本t检验, P<0.05,df=4), 而对裸甲藻的摄食率变化较小。双毛纺锤水蚤与已有研究中的锥形宽水蚤(Temora turbinata)[6]相类似, 弱的营养级联效应虽然释放了一部分裸甲藻对中肋骨条藻的摄食压力, 但是来自双毛纺锤水蚤较高直接摄食压力, 导致中肋骨条藻的生物量并没有增加, 反而是降低的, 也即[In(St/Sc)/t]是负值。双毛纺锤水蚤虽然是杂食性的, 但是在裸甲藻和中肋骨条藻两者间, 还是倾向于植食性为主, 因此在该食物链的营养传递过程中级联效应相对较弱, 双毛纺锤水蚤的直接摄食是中肋骨条藻最终生物量的决定因素。
实验室内的模拟实验采用的是纯的藻种和单一桡足类物种, 是一种理想化的食物链传递过程。天津近岸由于人为活动导致水体富营养化, 是赤潮频发的海域。赤潮发生时往往原因物种较单一, 如果存在微型浮游动物作为中间的摄食者, 我们可以认为跟实验室的模拟实验较类似。因此, 根据我们的实验结果推测, 在硅藻赤潮的初期时营养级联效应可能较明显, 而到赤潮的中期和后期级联效应会降低, 来自中型浮游动物的直接摄食是控制和缩短赤潮时间的重要因素。未来我们有必要对赤潮频发的硅藻和甲藻等的原因物种以及从群落水平, 更深入地研究其在食物网营养传递过程中作用和地位。
4 结论
本研究在实验室模拟了食物链双毛纺锤水蚤-裸甲藻-中肋骨条藻, 营养传递过程中级联效应介于0.018~0.12 d-1, 随中肋骨条藻生物量的增加而降低。级联效应的大小与双毛纺锤水蚤摄食行为的改变密切相关。双毛纺锤水蚤的摄食选择性与中肋骨条藻的平均生物量呈正相关, 与裸甲藻的平均生物量没有显著的相关性, 在中肋骨条藻低生物量时双毛纺锤水蚤偏向摄食裸甲藻; 在高生物量时选择摄食中肋骨条藻, 且对其摄食率显著高于摄食裸甲藻。双毛纺锤水蚤对中肋骨条藻的直接摄食高于营养级联效应, 决定了中肋骨条藻生物量的变化。
致谢: 感谢中国科学院海洋研究所MASCC海藻种质库为实验提供的藻种。
