低乳糖酸奶与普通酸奶的特性分析
2013-08-07徐爱才马成杰华宝珍杜昭平徐志平
徐爱才,马成杰,华宝珍,杜昭平,徐志平,刘 军
(1.光明乳业股份有限公司技术研究中心华中研究所,湖北武汉430040;2.武汉工业学院生物与制药工程学院,湖北武汉430023)
乳与乳制品是营养成分十分丰富的天然食品,是人类不可或缺的一类营养品。人刚出生时,肠道内乳糖酶活性最高,可以有效的降解乳及乳制品中的乳糖,但随着人的生长发育,体内乳糖酶活性呈规律性衰减,导致许多青少年、成年人和老年人存在着不同程度的乳糖不耐症,从而影响了人们对乳制品的正常摄入[1-4]。酸奶是牛乳经乳酸菌发酵后的一种乳制品,其牛乳中所含的乳糖在乳酸菌发酵后能降解约30%~40%[5],虽能一定程度上缓解乳糖不耐症,但对那些乳糖不耐症状严重的患者,尤其是老年人来说普通酸奶仍无法满足他们的需求,因此必须开发出低乳糖甚者无乳糖乳制品来满足市场的需求[6]。乳糖经乳糖酶分解后生成葡萄糖和半乳糖,不仅能有效解决乳糖不耐受的问题,同时赋予了牛乳更高的甜味,因此利用乳糖酶水解乳糖为低乳糖乳制品的开发提供了一种途径。鉴于此,本研究利用乳糖酶对牛乳中的乳糖进行酶解,并比较酶解前后乳酸菌的发酵曲线,发酵乳在贮藏过程中后酸化速度、黏度、活菌数量及主要风味物质含量的变化,以期为低乳糖酸奶的开发提供一定的技术支持。
1 材料与方法
1.1 材料与仪器
鲜牛乳 武汉光明乳品有限公司;发酵剂 丹麦DANISCO 公司;中性乳糖酶 哈尔滨华美生物技术股份有限公司;乳糖/半乳糖试剂盒 Boehringer Mannheim 公司;MRS Merck 公司 其它均为国产分析纯。
Brookfield Dv-1 VISCOM ETER 黏度仪 德国Marimex 公司;紫外可见分光光度计 日本岛津UV-1800;台式冷冻大容量高速离心机 德国eppendorf 公司;Cinac 发酵素酸化监控系统 法国ALLIANCE 公司;数显pH 计 瑞士梅特勒托利多公司;超净工作台 上海科学仪器厂;MIR-253 生化培养箱 日本SANYO 公司;SA-300VF 灭菌锅 德国Sturdy Industrial 公司。
1.2 实验方法
1.2.1 酸奶工艺流程 低乳糖酸奶工艺流程:鲜牛乳→预热(65~70℃) →加8%蔗糖→均质(140~160MPa) →杀菌(95℃,5min) →冷却(37~39℃) →酶解( 水解率70%) →灭酶(65℃,15min) →冷却至(42℃) 接入发酵剂发酵10h 后结束发酵。
普通酸奶工艺流程:鲜牛乳→预热(65~70℃) →加8%蔗糖→均质( 140~160MPa) →杀菌( 95℃,5min) →冷却(42℃) 接入发酵剂发酵10h 后结束发酵。
1.2.2乳糖水解率的测定 采用 Boehringer Mannheim 乳糖/半乳糖试剂盒测定乳糖的水解率,其中:水解率(%)=(1-水解后酸奶中的乳糖含量/水解前牛乳中的乳糖含量)×100%。
1.2.3 牛乳发酵过程中pH 的测定 采用Cinac 发酵素酸化监控系统对牛乳发酵过程中pH 的变化进行监测。
1.2.4 发酵乳酸度的测定 发酵乳的酸度以酚酞为指示剂,用0.1000mol/L 的NaOH 标准溶液滴定。滴定酸度以吉尔涅尔度(°T)表示,即每100mL 样品消耗NaOH 溶液的体积,每个实验重复3 次,取平均值[7]。
1.2.5 发酵乳黏度的测定 将发酵乳样品调温至20~22℃,采用Brookfield Dv-1 VISCOM ETER 黏度仪(MA)4#转子进行测定,其中转子转速为30r/min,测定时间为30s,重复测定3 次,取其平均值。
1.2.6 发酵乳中活菌数含量的测定 牛乳经发酵10h 后利用IKA RW20 digital 搅拌器600r/min 搅拌3min,分别在第0、5、10、15、20、25d 取样利用培养基MRS 采用稀释倾注平皿法检测发酵乳中乳杆菌的活菌数[8]。
1.2.7 发酵乳中双乙酰含量的测定 邻苯二胺可以与联二酮类反应生成2,3-二甲基并吡嗪,生成物的盐酸盐在335nm 波长下有最大吸收,可测出丁二酮的含量[9]。每个实验重复3 次,取平均值。
1.2.8 发酵乳中乙醛含量的测定 乙醛在酸性条件下与亚硫酸氢钠发生加成反应,其剩余的亚硫酸氢钠被碘氧化,在碱性条件下乙醛亚硫酸氢钠与碘定量反应,根据反应当量关系计算乙醛含量[10]。每个实验重复3 次,取平均值。
2 结果与分析
2.1 牛乳经乳糖酶水解前后发酵曲线的比较
牛乳经乳糖酶水解后巴氏杀菌,冷却至42℃按质量比0.02%添加发酵剂,同时以未经乳糖酶水解巴氏杀菌的牛乳做对照,利用Cinac 发酵素酸化监控系统对发酵过程中酸乳pH 变化进行监测,pH 变化如图1 所示。
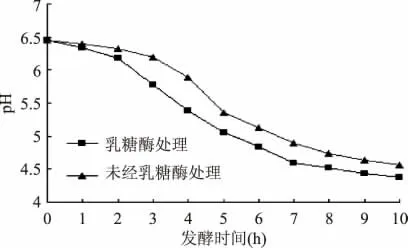
图1 样品pH-发酵时间的变化Fig.1 Changes of samples in pH during fermentation
由图1 可知,牛乳经乳糖酶水解后的发酵曲线与未经乳糖酶处理的发酵曲线有一定的差异。发酵10h 后,经乳糖酶处理的样品pH 由6.45 下降至4.38,而未经乳糖酶处理的样品的pH 由6.45 下降至4.56,发酵结束时前者的pH 比后者低0.18。接入发酵剂前1h,两种处理pH 下降均比较缓慢,这是由于菌体接入新环境后有一定的延迟期所致,2h 后经乳糖酶处理的样品pH 由6.45 下降至6.18,而未经乳糖酶处理的样品pH 由6.45 下降至6.32,经乳糖酶处理的样品的发酵速度比未经乳糖酶处理的样品的发酵速度略快。发酵2h 后,经乳糖酶处理的样品pH 急速下降,而未经乳糖酶处理的样品发酵3h 后pH 下降速率才有所增强。乳酸菌利用乳糖的过程可以分为两个阶段,第一个阶段是乳酸菌自身分泌少量的乳糖酶,将乳糖分解成葡萄糖和半乳糖;第二个阶段是乳酸菌利用葡萄糖或者半乳糖进行糖酵解[11]。在普通酸奶中,由于前期菌体数量较少,自身分泌的乳糖酶量较少;在低乳糖酸奶中,由于利用外源乳糖酶对乳糖进行水解,使得牛乳中存在大量的葡萄糖和半乳糖,而葡萄糖作为最直接能量供应单位而被乳酸菌直接利用,因此,与未经乳糖酶处理组相比,经乳糖酶处理的样品在发酵前期产酸更快,宏观表现为pH 下降更剧烈。
2.2 贮藏期间发酵乳酸度的变化
牛乳经发酵10h 后搅拌3min,4℃贮藏,低乳糖酸奶与普通酸奶的酸度在保质期内的变化如图2所示。
由图2 可知,在贮藏过程中,低乳糖酸奶和普通酸奶均有显著的后酸化,其中低乳糖酸奶的酸度由85°T 上升到112°T,上升了27°T,普通酸奶的酸度由76°T 上升到110°T,上升了34°T。虽然发酵结束时低乳糖酸奶的酸度比普通酸奶号高9°T,但在贮藏一段时间后酸度几乎相当,这与早期徐雅琴等研究的低乳糖酸奶数据相吻合[6]。酸乳在贮藏过程中,乳酸菌继续生长繁殖消耗残存的乳糖,半乳糖,葡萄糖,蔗糖产生乳酸导致酸度进一步升高。在贮藏前期,普通酸奶的后酸化速度比低乳糖酸奶快,这可能与乳酸菌生长至一定数量后自身乳糖酶分泌量增大,水解出足量葡萄糖加快了其新陈代谢的速度;而经乳糖酶处理的样品在葡萄糖逐渐耗尽后开始利用半乳糖和蔗糖新陈代谢放慢。低乳糖酸奶和普通酸奶在贮藏后期酸度逐渐趋于稳定,这可能与菌体处于衰退期自身产酸很弱,或部分菌体裂解后释放出的蛋白质被降解成碱性氨基酸有关[12]。

图2 样品在贮藏期间酸度的变化Fig.2 Changes of samples acidity in shelf life
2.3 贮藏期间发酵乳黏度的变化
图3 为低乳糖酸奶和普通酸奶在4℃贮藏条件下黏度变化情况。从图3 可以看出,在贮藏期间2 种酸奶的黏度显著增大,其中普通酸奶由469mPa·s 上升到628mPa·s,上升了159mPa·s,低乳糖酸奶由503mPa·s 上升到675mPa·s,上升了172mPa·s。酸乳黏度主要由蛋白质在酸性条件下发生变性沉淀和乳酸菌代谢产生的具有黏性的胞外多糖共同组成[13],是酸奶的一个重要指标,李全阳等通过对搅拌型酸奶流变学特性的研究认为,一般的酸奶测定黏度在400mPa·s 以上就具有较好的入口黏度[14-15],低乳糖酸奶在发酵终点时黏度比普通酸奶高34mPa·s,这可能与低乳糖酸奶中由于乳糖酶水解的作用,牛乳中葡萄糖含量较高,乳酸菌在前期利用葡萄糖产生更多的胞外多糖有关[16]。
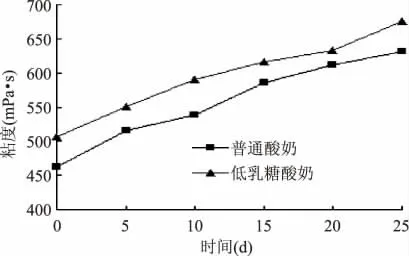
图3 样品在贮藏期间黏度的变化Fig.3 Changes of samples viscosity in shelf life
2.4 贮藏期间发酵乳中活菌含量的变化
低乳糖酸奶和普通酸奶在4℃贮藏过程中乳酸菌数量变化如图4 所示。从图4 可以看出,在贮藏期间,乳酸菌数量都有明显的先上升后下降趋势,低乳糖酸奶中乳酸菌数量略高于普通酸奶。低乳糖酸奶中乳酸菌数量在10d 后开始呈下降趋势,而普通酸奶在15d 后开始呈下降趋势,这可能与低乳糖酸奶中前期产生的乳酸较多,pH 较低有关。随着贮藏时间的延长,酸乳酸度的进一步提高,活菌数逐渐下降,25d 后普通酸奶中活菌数量已下降至1.4 ×108cfu/g,但活菌数仍高于国家标准的最低限(≥106cfu/mL)。酸乳在贮藏过程中,随着乳酸菌进一步的新陈代谢,乳酸含量逐渐升高,pH 逐渐下降,菌体的生长受到其代谢产物的抑制,同时菌株老化,死亡数增多,故逐渐出现活菌数量上的下降[17]。
2.5 贮藏期间发酵乳中双乙酰含量的变化
低乳糖酸奶和普通酸奶在4℃贮藏期间双乙酰含量变化见图5,由图5 可知,低乳糖酸奶中的双乙酰含量显著高于普通酸奶。其中,低乳糖酸奶中双乙酰含量由11.4mg/L 增加至13.6mg/L,普通酸奶中双乙酰含量由10.6mg/L 增加至12.7mg/L,酸乳中双乙酰含量都随时间的延长而升高,这亦说明酸奶随着贮藏时间的延长风味会更加浓厚,贮藏一段时间饮用风味更佳。乳酸菌发酵过程中,丁二酮由代谢中间产物α-乙酰乳酸经氧化脱羧产生,为初级代谢产物,因此在前期菌体代谢旺盛时合成较多,而在4℃冷藏期间增加量不大[18]。
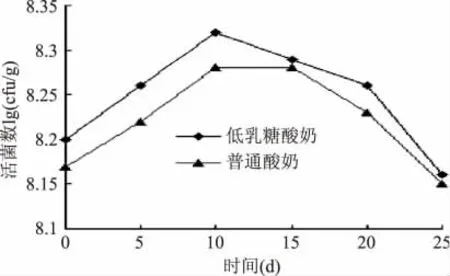
图4 样品在贮藏期间乳杆菌活菌数的变化Fig.4 Changes of samples viable cell numbers of Lactobacillus in shelf life
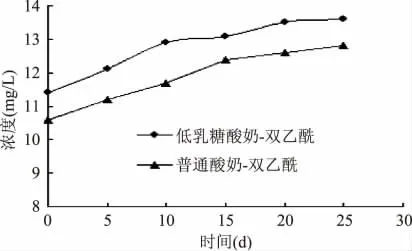
图5 样品在贮藏期间双乙酰含量的变化Fig.5 Changes of samples contend of diacetyl in shelf life
2.6 贮藏期间发酵乳中乙醛含量的变化
图6 为低乳糖酸奶和普通酸奶在4℃贮藏条件下随时间延长乙醛含量变化情况,由图6 可知,随着时间的延长,低乳糖酸奶与普通酸奶中乙醛的含量均显著提高,其中低乳糖酸奶中乙醛含量由17.1mg/L 上升到20.5mg/L,普通酸奶中乙醛含量由16.5mg/L 上升到19.6mg/L,低乳糖酸奶增幅略高于普通酸奶。在酸乳发酵期间,乙醛由丙酮酸脱羧酶或丙酮酸氧化酶催化直接产生,也可先由丙酮酸脱氢酶催化生成中间产物乙酰辅酶A 间接产生;低乳糖酸奶在前期发酵产生更多的中间产物,因此乙醛含量较普通酸奶更高[18-19]。
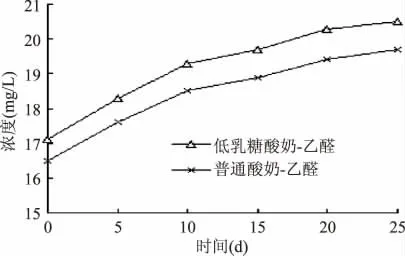
图6 样品在贮藏期间乙醛含量的变化Fig.6 Changes of samples contend of acetaldehyde in shelf life
3 结论
本文对低乳糖酸奶与普通酸奶在发酵过程中pH 的变化及发酵乳在4℃贮藏期间酸度、黏度、活菌数含量、主要风味物质双乙酰和乙醛变化进行了比较,结果表明低乳糖酸奶在发酵过程中发酵更快,贮藏过程中酸度变化更小,而且黏度、活菌数、风味物质含量均高于普通酸奶。低乳糖酸奶与普通酸奶相比具有更高的品质,更具市场前景。
乳糖不耐症一直影响着消费者对牛乳及乳制品的需求。虽然近几年随着乳业技术的提升,低乳糖巴氏奶、低乳糖UHT 奶、低乳糖奶粉相继被开发出来并逐渐被消费者接受,但其乳糖的含量仍无法满足乳糖不耐症严重的人群;低乳糖酸奶通过乳糖酶与乳酸菌发酵相结合进一步降低了酸奶中乳糖的含量,丰富了乳制品市场,相信随着乳业技术的发展,乳糖不耐受将会得到根本性解决。
[1]Davidson G P,Goodwin D,Robb T A.Incidence and duration of lactose malabsorption in children hospitalized with acute enteritis: study in a well-nourished urban population[J].The Journal of Pediatrics,1984,105(4) :587-590.
[2] Nagaraj M,Sharanagouda B,Manjunath H,et al.Standardization of different levels of lactose hydrolysis in the preparation of lactose hydrolyzed yogurt[J].Iranian Journal of Veterinary Research,Shiraz University,2009,10(2) :132-136.
[3]Jeanette N K,Jill N,Alan R,et al.The prevalence of selfreported Lactose Intolerance and the consumption of dairy foods among african american adults are less than expected[J].Journal of the National Medical Association,2011,103(1) :36-45.
[4]Daniel L,Swagerty Jr,Anne D,et al.Lactose intolerance[J].American Family Physician,2002,65(9) :1845-1850.
[5]Ibarra A,Acha R,Callahan M T,et al.Optimization and shelf life of a low-lactose yogurt with Lactobacillus rhamnosus HN001[J].Journal of Dairy Science,2012,95(7) :3536-3548.
[6]徐雅琴,冯大伟.中性乳糖酶Maxilact 对酸奶感官品质和发酵过程的影响.中国食品添加剂[J],2009,1:81-84.
[7]张水华.食品分析[M].北京:中国轻工业出版社,2004.
[8]凌代文.乳酸菌分类鉴定及实验方法[M].北京:中国轻工业出版社,1999.
[9]吕嘉枥,李成涛.酸奶中双乙酰含量的测定方法初探[J].中国乳品工业,2006,34(1) :21-22.
[10]李妍,邢慧敏,邵亚东,等.发酵乳中丁二酮和乙醛含量检测方法探讨[J].分析与检测,2008,34(3) :157-159.
[11]郭本恒.益生菌[M].北京:化学工业出版社,2004.
[12]郭清泉,张兰威,林淑英.酸奶发酵机理及后酸化控制措施[J].食品与发酵工业,2001,27(2) :80-83.
[13] Skriver A,Roemer R H,Qvist K B. Rheological characterization of stirred yoghurt: viscometry[J]. Journal of Texture Study,1993,24:185-190.
[14]李全阳,夏文水.酸乳中乳酸菌所产胞外多糖特性的初步研究[J].食品科学,2009,25(2) :80-84.
[15]徐成勇,郭本恒,郑思聪,等.弱后酸化酸奶发酵剂的筛选[J].中国乳品工业,2007,35(3) :12-16.
[16]Andre R M,Ricardo L M,Janaina F M,et al.Simultaneous enzymatic hydrolysis and lactic fermentation to obtain a yogurt with lactose content[J].Food Scinece and Technology,2012,36(5) :551-559.
[17]尚玉琳,孟祥晨.不同酸奶发酵剂的发酵性能比较[J].食品科技,2011,36(10) :41-45.
[18]Hugenholtz J,Sybesma W,Nierop G M,et al.Metabolic engineering of Lactic acid bacteria for the production of nutraceuticals[J].Antonie Leeuwenhoek,2002,82:217-235.
[19]韩希妍,孙大庆,相丽,等.乳酸菌风味代谢物质的基因调控[J].微生物学报,2007,47(6) :1105-1109.
