叶面喷施胺鲜酯及其与油菜素内酯复配液对油用牡丹生理特性及产量的影响
2024-04-01李晓鹏肖瑞雪张利霞张亚冰余世玺陈轲李仁杰罗宁侯小改
李晓鹏 肖瑞雪 张利霞 张亚冰 余世玺 陈轲 李仁杰 罗宁 侯小改
摘要:为筛选出适合油用牡丹高产高效栽培中的植物生长调节剂应用技术,本试验以凤丹为材料,分别于盛花期(4 月7 日)、籽粒形成期(4 月18 日)、果荚与籽粒快速生长期(5 月10 日)叶面喷施不同质量浓度的胺鲜酯(D1、D2 和D3)及其与油菜素内酯复配液(BD1、BD2 和BD3),研究其对植株生理特性、籽粒产量指标(百粒重、单果籽粒重、单株籽粒重及籽粒产量)的影响。 结果表明,与对照组相比,喷施植物生长调节剂处理的牡丹叶片生理特性和籽粒产量关键参数均有明显提高:7 月3 日时,BD2 处理叶片SOD 活性较对照显著提高5.24%;5 月19 日和7 月3 日时,所有处理叶片MDA 含量均较对照显著降低;7 月3 日时,所有处理叶片可溶性糖含量与对照均有显著性差异,其中BD2 处理最高,较对照增加33.84%;6 月10 日时,所有处理叶片总叶绿素含量均显著高于对照,其中BD3 处理增幅最大,为98.11%;3 个测定时期,除6 月10 日的BD3 处理外其他处理叶片净光合速率均显著高于对照,且随着胺鲜酯质量浓度的提高呈现先增加后降低趋势。 所有处理单株籽粒重及籽粒产量均显著高于对照,其中BD3 增幅均最大,分别为10.45%与10.44%。 相关性分析表明,SOD 活性与Chl a、Chl b、Car、总叶绿素含量和净光合速率呈极显著正相关,与可溶性糖含量呈显著正相关,与MDA 含量呈极显著负相关;总叶绿素含量与净光合速率呈极显著正相关,净光合速率与单果籽粒重、单株籽粒重和籽粒产量呈显著正相关。 综之,12、16 mg/ L 胺鲜酯与0.05 mg/ L 油菜素内酯复配喷施油用牡丹植株的生理特性表现优异,并可获得较高籽粒产量。
关键词:油用牡丹;凤丹;胺鲜酯;油菜素内酯;生理特性;籽粒产量
中图分类号:S565.9文献标识号:A文章编号:1001-4942(2024)01-0097-08
凤丹(Paeonia ostii ‘Fengdan )属于芍药科(Paeoniaceae) 芍药属(Paeonia) 牡丹组木本植物,是我国的十大传统名花之一,不仅具有较高的观赏及药用价值,还是一种重要的油料作物[1] 。作为重要的油用牡丹品种,凤丹籽油含有多种人体必需的不饱和脂肪酸(α-亚麻酸、油酸、亚油酸等),约占总脂肪酸的92.1%,具有增强免疫力、提高记忆力、降血糖、降血脂、保护肝脏、抗肿瘤和抗氧化衰老等重要作用,并且其中可参与细胞膜和生物酶合成的α-亚麻酸高达67%左右[2] 。 然而,凤丹籽粒产量较低,限制农户种植的积极性,进一步影响籽油开发。 鉴于消费者对食用油品质和性价比的双重需求,解决凤丹籽粒产量低的问题是提升凤丹籽油核心竞争力的关键。
油用牡丹凤丹在实际生产中,由于缺乏配套的高产栽培管理技术等原因,导致产量严重偏低,故而如何提高产量,成为解决凤丹种植率低的关键[3] 。 长期以来,国内外学者就如何通过改善栽培措施提高作物产量潜力开展过广泛研究,其中有关植物生长调节剂对粮食作物产量和品质影响方面的研究较多,而关于叶面喷施植物生长调节剂对油用牡丹生理特性及籽粒产量影响的研究较少。 前人研究发现,适宜浓度脱落酸能改善油用牡丹在夏季高温条件下的生理特性,提高植株光合能力,有助于提高油用牡丹的抗逆性[4] ;外源油菜素内酯能显著提高牡丹叶片抗氧化能力,提高凤丹光合能力[5] 。
胺鲜酯(diethyl aminoethyl hexanoate,DA-6)作为一种植物生长调节剂,能够促进植物生长发育,促进蔗糖代谢与光系统中关键酶基因的表达,提高植株抗氧化酶活性与光合色素捕获光能的转化率,加快光合进程;还能调节养分平衡,提高碳氮代谢比率,促进根系发育,从而提高植株的抗逆性和产量,在植物生长与产量调控方面具有重要的应用前景[6-9] 。 油菜素内酯(brassinolide,BRs)可以通过提高植物体内SOD 活性,增强其同功酶的表达,清除细胞在胁迫环境中积累的自由基,稳定膜结构,使电解质外渗率下降,降低对叶绿素的破坏作用,从而改善植物品质[10] 。 目前,关于外源植物生长调节剂对油用牡丹凤丹生理特性及籽粒产量影响的研究尚未见报道。 基于此,本试验通过研究叶面喷施胺鲜酯及其与油菜素内酯复配液对油用牡丹凤丹生理特性及产量的影响,旨在为进一步完善油用牡丹高产高效栽培技术体系提供理论支撑。
1 材料与方法
1.1 试验地概况
试验在河南科技大学牡丹试验基地(112°36′19.65″E,34°39′55.43″N)进行。 试验地土壤为黄潮土,pH 值为7.12,0~40 cm 土层含有机质12.98g/ kg、全氮0.81 g/ kg、碱解氮73.32 mg/ kg、有效磷12.01 mg/ kg 和速效钾146.25 mg/ kg。 该地位于暖温带,四季分明,夏季平均最高温度为31 ℃,冬季平均最低温度为1 ℃,年均夜间温度为11 ℃,白天平均温度为21 ℃,年均降水量为578.2 mm,年均日照时数为1 791.6 h,海拔136 m。
1.2 试验设计
每小区选取20 株长势均匀一致、无病虫害的七年生鳳丹(株行距为40 cm×60 cm)进行试验。小区间挖置宽为45 cm 的沟,并设置宽为1 m 的保护行,以减小边际效应的影响,提高试验准确度。 试验采用随机区组设计,以喷施清水为对照,共设置7 个处理,重复5 次。 其中,单独处理:胺鲜酯质量浓度梯度为:0、8、12、16 mg/ L,依次用CK、D1、D2 和D3 表示;复配处理:将0.05 mg/ L油菜素内酯分别溶于三种质量浓度胺鲜酯溶液,依次用BD1、BD2 和BD3 表示。 处理方法:分别于盛花期(4 月7 日)、籽粒形成期(4 月18 日)、果荚与籽粒快速生长期(5 月10 日)对整株正反叶面进行喷施,喷施量为每株150 mL。 按照凤丹生长发育规律,分别于2019 年5 月19 日、6 月10日和7 月3 日选取植株倒三叶用于生理指标测定。
1.3 测定指标及方法
采用氮蓝四唑法测定超氧化物岐化酶(SOD)活性[11] ;采用硫代巴比妥酸法测定丙二醛含量[11] ;采用蒽酮比色法测定可溶性糖含量[11] ;采用80%丙酮浸泡法测定光合色素含量[11] ;采用LI-6400 便携式光合作用系统测定净光合速率(Pn)。
待果荚成熟后,即呈蟹黄色并轻微开裂时(7月25 日) 采集果荚,自然阴干后测定单果籽粒重、百粒重、单株籽粒重和籽粒产量。
1.4 数据处理与分析
采用Microsoft Excel 2010 进行数据统计与图表制作,所有数据以平均值±标准差(mean±SD)表示。 采用SPSS 16.0 软件对数据进行单因素(one-way ANOVA)方差分析、Duncans 多重比较及相关性分析。
2 结果与分析
2.1 不同处理下凤丹叶片生理指标的变化
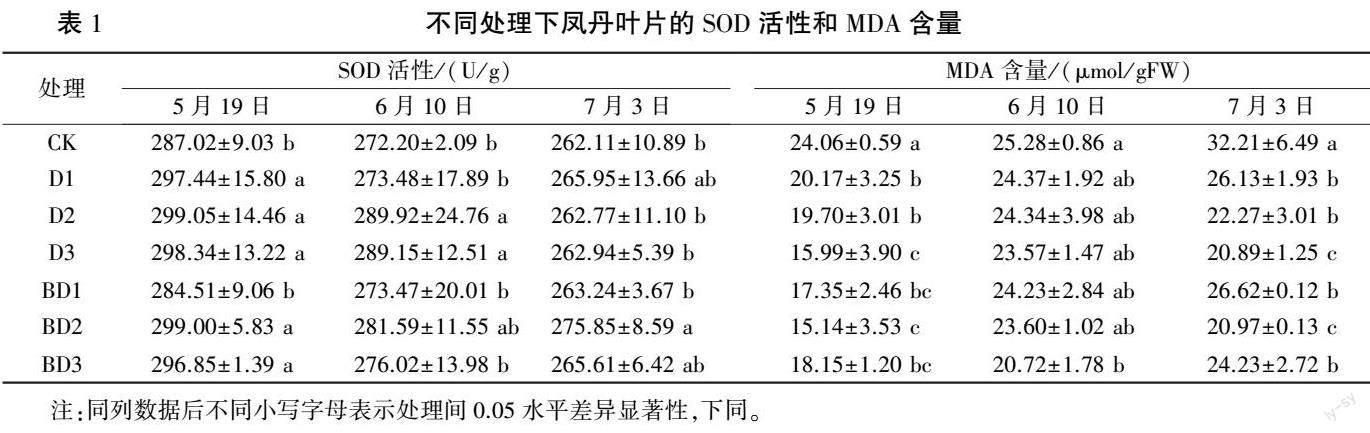
2.1.1 凤丹叶片SOD 活性和MDA 含量的变化 由表1 可知,5 月19 日,除BD1 处理叶片SOD 活性与对照(CK)无显著差异外,其他处理均显著升高;D、BD 处理组SOD 活性随着胺鲜酯质量浓度的升高均呈现先升高后降低趋势。 6 月10 日,D2、D3 处理叶片SOD 活性显著高于对照,其他处理与对照差异不显著;D、BD 处理组叶片SOD 活性随着胺鲜酯质量浓度的升高基本呈先升高后降低趋势。 7 月3 日,BD2 处理叶片SOD 活性显著高于对照,增幅为5.24%,其他处理与对照无显著差异;BD 处理组SOD 活性随着胺鲜酯质量浓度的升高呈先升高后降低趋势。
由表1 可知,5 月19 日,所有处理凤丹叶片MDA 含量均显著低于对照;BD 处理组随胺鲜酯质量浓度升高呈先降低后升高趋势,BD2 处理MDA 含量最低,较对照降37.08%。 6 月10 日,BD3 处理叶片MDA 含量与对照相比显著降低,其他处理均低于对照但无显著差异。 7 月3 日,所有处理叶片MDA 含量均显著低于对照,D3、BD2 处理降幅尤其显著, 分别降35. 15% 和34.90%。
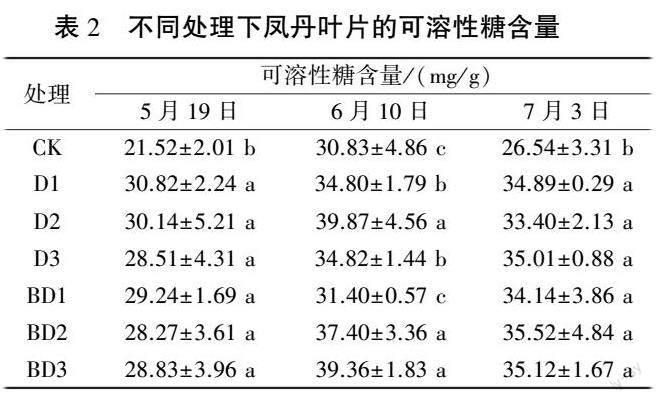
2.1.2 凤丹叶片可溶性糖含量的变化 由表2 可知,5 月19 日,所有处理与对照相比,可溶性糖含量均显著提高。 6 月10 日,除BD1 外,其他处理均与对照有显著性差异,其中D2 和BD3 可溶性糖含量较高,较对照增幅分别为29.32%和27.67%。 7 月3日,所有处理均显著高于对照,其中BD2 处理可溶性糖含量最高,较对照增加33.84%。
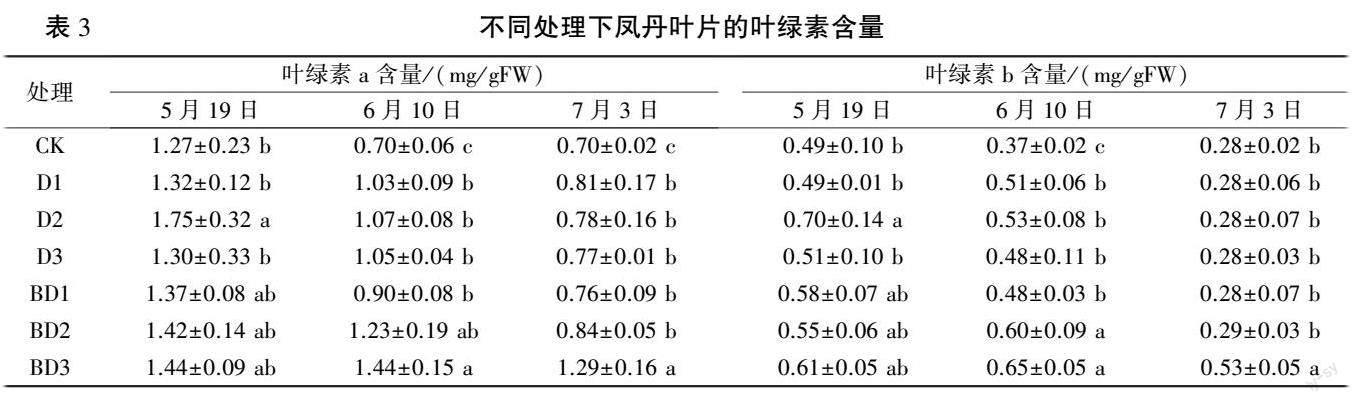
2.1.3 凤丹叶片光合色素含量的变化 由表3可以看出,5 月19 日,D2 处理叶绿素a(Chl a)含量达1.75 mg/ g,比对照显著增加37.80%。 6 月10 日和7 月3 日,所有处理叶片叶绿素a 含量均显著高于对照。
5 月19 日与6 月10 日,D 处理组叶片叶绿素b(Chl b)含量随胺鲜酯质量浓度的升高呈先升高后降低趋势,并均在D2 处理下显著高于对照,比对照分别增加42.86%和43.24%。 6 月10 日,随胺鲜酯质量浓度的升高,BD 处理组叶绿素b 含量呈逐渐增加趋势,并均显著高于对照,BD3 处理增加75.68%。 7 月3 日,BD3 处理与对照相比叶绿素含量显著提高,增幅为89.89%,其余处理与对照相比无显著差异。
5 月19 日,仅D2 处理叶片类胡萝卜素(Car)含量显著高于对照(36.03%),其余处理均无显著差异(表4)。 6 月10 日,所有处理叶片类胡萝卜素含量均显著高于对照,其中BD2 处理增幅最大,为93.33%。 7 月3 日,所有处理叶片类胡萝卜素含量均显著高于对照。
5 月19 日,D2 处理叶片总叶绿素含量显著高于对照,增幅达40.16%,其余处理均高于对照但无显著差异(表4)。 6 月10 日,所有处理叶片总叶绿素含量均显著高于对照,其中BD3 处理增幅最大,为98.11%。 7 月3 日,除BD3 处理显著高于对照外,其余处理均高于对照但差异不显著。
2.1.4 凤丹叶片净光合速率的變化 由表5 可知,5 月19 日,所有处理叶片净光合速率(Pn)均显著高于对照,且BD 处理组随着胺鲜酯质量浓度的升高呈现先增加后降低趋势,BD2 处理对Pn的提升作用最佳,较对照增加35.17%。 6 月10日,D、BD 处理组随着胺鲜酯质量浓度的升高,叶片净光合速率均呈现先增加后降低趋势。 7 月3日,所有喷施处理叶片净光合速率均显著高于对照,且随着胺鲜酯质量浓度的升高均呈现先增加后降低趋势,其中D2 处理较对照增幅最大,为75.69%。
2.2 不同处理下凤丹籽粒产量指标的变化
从百粒重数据(表6)看,D1 处理显著大于对照,其他处理与对照无显著差异。 各处理单果籽粒重均显著高于对照,其中BD2 处理单果籽粒重达到最大值,较对照增幅为46.92%。 单株籽粒重与籽粒产量相似,所有处理均显著高于对照,其中BD3 增幅最大,分别为10.45%与10.44%。
2.3 不同处理下凤丹生理特性与籽粒产量指标的相关性分析
由表7 可知,SOD 活性与Chl a、Chl b、Car、总叶绿素含量和净光合速率呈极显著正相关,与可溶性糖含量呈显著正相关,与MDA 含量呈极显著负相关。 MDA 含量与可溶性糖、Chl a、Chl b、Car、总叶绿素含量和净光合速率呈极显著负相关。 可溶性糖含量与Chl a、Chl b 和总叶绿素含量呈显著正相关,与Car 和净光合速率呈极显著正相关;Chl a 与Chl b、Car、总叶绿素含量和净光合速率呈极显著正相关;Chl b 与Car、总叶绿素含量呈极显著正相关,与净光合速率呈显著正相关;Car 与总叶绿素含量、净光合速率呈极显著正相关;总叶绿素含量与净光合速率呈极显著正相关;净光合速率与单果籽粒重、单株籽粒重和籽粒产量呈显著正相关。
3 讨论
3.1 不同处理对凤丹生理特性的影响
本研究表明,5 月19 日,除BD1 处理与对照无显著差异外,其他处理凤丹叶片SOD 活性与对照相比均显著提高;6 月10 日,D2、D3 处理SOD活性显著高于对照,D、BD 处理组SOD 活性随着胺鲜酯质量浓度的升高均呈先升高后降低趋势;7月3 日,BD2 处理显著高于对照,增幅为5.24%。以上说明适宜质量浓度的胺鲜酯及其与油菜素内酯复配喷施能对SOD 活性产生显著促进作用,增加活性氧的清除能力。 这与任增斌等[12] 的研究结果一致。 5 月19 日和7 月3 日,所有处理的MDA 含量均显著低于对照,这说明适宜质量浓度的胺鲜酯单施及复配处理均显著降低MDA 含量,保护细胞膜系统结构的完整性,维持植株的正常生长发育。 胺鲜酯可通过影响植物内源激素的水平来调节植物体内的酶活性[13] ,外源BR 处理可调节逆境环境下植物体内的代谢平衡[14] ,两种物质协同作用,能够及时清除活性氧和降低膜脂过氧化程度以提高凤丹抗氧化能力,并延缓植株衰老[15] 。 由此说明,胺鲜酯与油菜素内酯两种植物生长调节剂具有较好的协同作用,有助于降低植株细胞膜过氧化水平并提高抗氧化酶活性。
本研究表明,5 月19 日与7 月3 日,所有处理凤丹叶片可溶性糖含量较对照都有显著增加,即胺鲜酯处理后凤丹叶片可溶性糖含量提高,这与闻祥成[16] 、彭静[17] 等的研究结果一致。 其原因可能是,胺鲜酯能够提高凤丹叶片中蔗糖磷酸合成酶和蔗糖合成酶活性以促进可溶性糖的合成[18] ,或通过提高RuBPCase 和PEPCase 活性从而促进植物体内可溶性糖积累,进而改善植物的光合生理特性[19] 。 BR 能够提高植物中蔗糖磷酸合成酶活性,在提高植物蔗糖积累量方面具有积极作用[20] ,同时促进蔗糖等有机物质更多地向果实转移,最终提高植物产量[14] 。
本试验条件下,6 月10 日所有处理凤丹叶片的总叶绿素与Car 含量均显著高于对照;除6 月10 日的BD3 处理外,5 月19 日、6 月10 日与7 月3 日其他处理叶片Pn 均显著高于对照。 这表明单施及复配处理组能够通过调节凤丹植株体内生理代谢过程,启动抗氧化酶系统,提高叶片渗透调节物质含量,降低植株体内渗透势,维持渗透平衡,继而提高凤丹植株的抗氧化能力,减缓叶绿素降解速度,保护叶绿体结构的完整性,提高凤丹叶片光合色素含量,延缓衰老进程,提升植株的光合能力。 这与朱凤荣等[21] 的研究结果一致。 外源喷施植物生长调节物质必然会影响凤丹体内某些激素间的平衡,而这种关系的改变必然引起其生理的甚至形态上的效应[22] 。 生理指标相关性分析表明,SOD 活性与Chl a、Chl b、Car、总叶绿素、可溶性糖含量和Pn 互为正相关;MDA 含量与可溶性糖、Chl a、Chl b、Car、总叶绿素含量和净光合速率呈极显著负相关。 净光合速率与其他指标的相关顺序为:单果籽粒重>SOD 活性>Chl a>Car>可溶性糖含量>总叶绿素含量>Chl b>单株籽粒重。 以上说明适宜质量浓度DA-6 单施及与BR复配喷施,凤丹叶片的抗氧化酶活性增加,渗透调节能力增强,叶绿素含量增加,光合作用能力进一步增强,有利于产量的正向积累。
3.2 不同处理对凤丹籽粒产量的影响
本研究表明,BD 处理组单果籽粒重显著高于对照,说明外源DA-6 与BR 配施能够显著增加油用牡丹单果籽粒重。 聂乐兴[23] 采用10 mg/ LDA-6 溶液处理玉米增产达10.0%,进一步证实DA-6 对作物具有明显的增产作用。 外源BR 处理可通过提高植物叶片的叶绿素含量、净光合速率、光化学效率以及碳酸酐酶、Rubisco 活性来抑制高光强对光化学反应中心的损伤,缓解不良环境对光合作用原初反应的抑制和对光合电子传递过程的阻碍,促进植物光合机构对CO2 的利用效率,增强植物叶片的光合碳同化能力和光适应状态下PSⅡ进行光化学反应的能力,保护凤丹的光合器官与结构[24-25] 。 模式植物拟南芥经外源BR处理后籽粒重显著增加[26] ,表明BR 与DA-6 相似,均具有促进籽粒增产的作用。 本研究中,BD處理显著改善凤丹叶片的光合功能,单果籽粒重显著增加,促进同化产物向籽粒的供给。 说明胺鲜酯与BR 复配处理发挥出增效作用,使籽粒产量明显增加。 这与李蒙等[27] 采用0.1 mg/ L EBR处理樱桃番茄后单株总产量增加31.13%和郑强卿等[28] 采用30 mg/ L DA-6 与水杨酸、外源BR复配喷施骏枣增产25.18%的研究结果一致。 叶绿素含量与净光合速率呈极显著正相关,净光合速率与单果籽粒重、单株籽粒重和籽粒产量呈显著正相关。 单株籽粒重作为衡量籽粒产量高低的重要指标之一,一定程度上能够准确评价产量高低;植株净光合速率提高,说明光合产物积累增加,更有利于果实的正向积累[29] 。
4 结论
本研究表明,适宜质量浓度的胺鲜酯及其与油菜素内酯配合喷施能够增强油用牡丹植株的生理特性与光合能力,并能够增加油用牡丹籽粒产量,尤其是BD2 与BD3 处理(12、16 mg/ L 胺鲜酯与0.05 mg/ L 油菜素内酯复配)能够显著提高油用牡丹籽粒产量。 本研究结果可为油用牡丹凤丹高产高效栽培中的植物生长调节剂应用提供一定参考。 此外,本研究是小区试验结果,或许与大田实际栽培有一定差异,因此尚需在大田实际生产与栽培过程中进一步验证。
参 考 文 献:
[1] Duan X G, Liu W, Wang X J, et al. Effects of phosphorus fer ̄tilization on growth characteristics,fatty acid composition,andseed yields of Fengdan(Paeonia ostii T. Hong & J. X. Zhang)[J]. Hort. Science,2022,57(6):733-740.
[2] Liu W, Yin D X, Zhang T, et al. Major fatty acid compositionsand antioxidant activity of cultivated Paeoniaostii under differ ̄ent nitrogen fertilizer application[J]. Chemistry Biodiversity,2020,17(12):e2000617.
[3] 馬会萍,彭正锋,冀含乐,等. 油用牡丹产业化的制约因素及对策[J]. 安徽农学通报, 2021,27(8):51-53.
[4] 肖瑞雪,王晓静,张利霞,等. 外源脱落酸对油用牡丹‘凤丹生理特性的影响[J]. 基因组学与应用生物学,2020,39(6):2705-2711.
[5] 肖瑞雪,吕静霞,贾长松,等. 外源油菜素内酯对油用牡丹‘凤丹生理特性的影响[J]. 植物生理学报,2018,54(9):1417-1425.
[6] 李合生. 现代植物生理学[M]. 第3 版. 北京:高等教育出版社,2012.
[7] James H, Keithl J H, Yokoyama H. Regulation of crop growthand yield by tertiary amine bioregulators[M] / / Gausman H W.Plant Biochemical Regulator,New York,1991:223-246.
[8] 关朋霄. DA-6 提高番茄幼苗低夜温抗性途径的研究[D].沈阳:沈阳农业大学,2020.
[9] 郝青南,汪嫒嫒,龙泽福,等. DA-6 对南方大豆品种性状、产量和品质的影响[J]. 大豆科学,2021,40(6):799-804.
[10] 曹云英,赵华. 高温胁迫下油菜素内酯对水稻幼苗的保护作用[J]. 中国水稻科学, 2007,21(5):525-529.
[11] 闫敏,王艳,鲍荆凯,等. 混合盐碱胁迫对骏枣渗透调节物质和抗氧化酶活性的影响[J]. 山东农业科学,2022,54(5):37-43.
[12] 任增斌,陈法志,舒常庆,等. 外源表油菜素内酯对牡丹耐热性的影响[J]. 江汉大学学报(自然科学版),2018,46(5):446-453.
[13] 刘笑鸣,顾万荣,李从锋,等. 化学调控和氮肥对高密度下春玉米光热水利用效率和产量的影响[J]. 中国农业科学,2020,53(15):3083-3094.
[14] Wei L J, Deng X G, Zhu T, et al. Ethylene is involved inbrassinosteroids induced alternative respiratory pathway in cu ̄cumber(Cucumis sativus L.) seedlings response to abiotic stress[J]. Frontiers in Plant Science,2015,6:982.
[15] Ali B, Hasan S A, Hayat S, et al. A role for brassinosteroidsin the amelioration of aluminium stress through antioxidant sys ̄tem in mung bean(Vigna radiata L. Wilczek) [J]. Environ ̄mental and Experimental Botany,2008,62(2):153-159.
[16] 闻祥成,田华,潘圣刚,等. 叶面喷施植物生长调节剂对水稻产量及叶片保护酶活性的影响[J]. 西南农业学报,2015,28(2):550-555.
[17] 彭静,杨雪,罗梦娜,等. 喷施组合型生长调节剂对不同品种小麦冬前分蘖和生长及抗寒性的影响[J]. 干旱地区农业研究,2019,37(1):137-143.
[18] 伍宝朵,周蓉,陈海峰,等. 蔗糖载体调控作物“源、库”分配的研究进展[J]. 中国农学通报,2012,28(6):19-23.
[19] 聂乐兴. 四种植物生长调节剂对超高产玉米的调控作用研究[D]. 泰安:山东农业大学, 2010.
[20] 于安玲. 芸薹素(BR)和赤霉素(GA) 对小麦籽粒淀粉积累、粒度分布及加工特性的影响[D]. 泰安:山东农业大学,2011.
[21] 朱凤荣,邱宗波. 种植密度和植物生长调节剂对小麦衰老和产量构成的影响[J]. 河南农业科学,2004(8):18-21.
[22] Tang Y L,Wen X G,Lu C M. Differential changes in degrada ̄tion of chlorophyll ̄protein complexes of photosystem I and phot ̄osystem Ⅱ during flag leaf senescence of rice[J]. Plant Physi ̄ology and Biochemistry,2005,43(2): 193-201.
[23] 聂乐兴. 四种植物生长调节剂对超高产玉米的调控作用研究[D]. 泰安:山东农业大学, 2010.
[24] Andersson I, Backlund A. Structure and function of Rubisco[J]. Plant Physiology and Biochemistry,2008,46(3):275-291.
[25] Yamori W, Suzuki K, Noguchi K O, et al. Effects of Rubiscokinetics and Rubisco activation state on the temperature de ̄pendence of the photosynthetic rate in spinach leaves from con ̄trasting growth temperatures[J]. Plant Cell and Environment,2006,29(8):1659-1670.
[26] Huang H Y, Jiang W B, Hu Y W, et al. BR signal influencesArabidopsis ovule and seed number through regulating relatedgenes expression by BZR1[J]. Molecular Plant,2013,6(2):456-469.
[27] 李蒙,束胜,郭世荣,等. 24-表油菜素内酯对樱桃番茄光合特性和果实品质的影响[J]. 西北植物学报,2015,35(1):138-145.
[28] 郑强卿,陈奇凌,王晶晶,等. 不同植物生长调节剂对骏枣内源激素与果实品质的影响[J]. 新疆农业科学,2019,56(3):430-437.
[29] 王春丽,海江波,田建华,等. 油菜终花后角果和叶片光合对籽粒产量和品质的影响[J]. 西北植物学报,2014,34(8):1620-1626.
