镉胁迫对黄秋英生理及富集特性的影响
2024-04-01李源恒赵春莉郭宏亮陈元晖刘翰升王晟旭
李源恒 赵春莉 郭宏亮 陈元晖 刘翰升 王晟旭
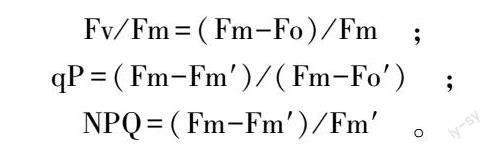

摘要:为给土壤镉(Cd)污染地区提供具有景观修复性的植物修复材料,本试验采用盆栽方法,研究不同质量浓度镉(0、0.3、3、30、60、120、180 mg/ kg)对黄秋英(Cosmos sulphureus)植株生理及富集特性的影响。 结果表明:胁迫15 d 时,随着镉浓度增加,黄秋英叶片可溶性蛋白、叶绿素a、叶绿素b、类胡萝卜素含量和SOD 活性均呈先升后降趋势,MDA 含量逐渐下降,POD 活性先降后升;ETR、qP、Tr 在镉浓度0.3 mg/ kg 时达到最高,Pn 与Gs 在镉浓度3 mg/ kg 时达到最高,NPQ 在镉浓度30 mg/ kg 时降到最低,Fv/ Fm 在镉浓度60 mg/ kg 时达到最高,Fo 在镉浓度180 mg/ kg 时降到最低。 胁迫30 d 时,其可溶性蛋白含量、SOD 活性先升后降,MDA 含量先升后降再升高,POD 活性先降后升再下降,叶绿素a、叶绿素b、类胡萝卜素含量先降后升;镉浓度0.3mg/ kg时Fv/ Fm 达到最高,镉浓度3 mg/ kg 时NPQ 达到最高,而Tr、Pn、Gs 降到最低,镉浓度30 mg/ kg 时qP达到最高、Fv/ Fm 降到最低,镉浓度60 mg/ kg 时Pn、Gs 达到最高,而Fo、NPQ 降到最低,镉浓度120 mg/ kg 时Ci 降到最低,镉浓度180 mg/ kg 时Tr、ETR 达到最高。 镉浓度<180 mg/ kg 时,黄秋英各器官镉含量表现为茎>根>叶;镉浓度≥180 mg/ kg 时为根>茎>叶。 镉浓度≥120 mg/ kg 时黄秋英根、茎及地上部镉含量均超过镉超富集植物的临界值。 综上看出,黄秋英在高浓度镉胁迫下具有较高的耐受力,同时具备较强的镉富集能力,可作为具有景观修复性的植物修复材料。
关键词:黄秋英;镉胁迫;光合生理;光响应;叶绿素荧光;富集特性
中图分类号:S567.21文献标识号:A文章编号:1001-4942(2024)01-0081-10
我国经济飞速发展的同时伴随着农工业资源的过度开发与超前利用,大量农工业活动对相应区域出现的重金属污染负有主要责任[1] ,它严重危害当地居民的生存环境、生命健康、生活质量。土壤镉污染经调查已是我國无机物污染之首[2] ,位点超标率达7.0%且远超自然背景值[3] 。 土壤中的镉可以通过呼吸及与皮肤接触等直接进入人体,也可通过富集作用经食物链间接进入人体,对人体造成肝肾功能损坏、结石和贫血等危害,严重影响我国环境安全[4-5] ,因此对镉污染的防范与治理已然成为当下亟待解决的课题之一。
植物修复技术是重金属污染修复方法中成本低、易操作、无二次污染并同时能够美化环境的友好型解决方案[6] ,它通过植物的生命活动将污染土壤中的重金属富集或降解,从而达到修复和治理的效果[7] 。 在植物修复实际应用中,通常选择富集能力较强的植物,而菊科植物在修复土壤镉污染中具有重要作用[8] :鬼针草(Bidens pilosa)镉富集系数达到4.16,富集效果良好[9] ;野外调查发现,超富集植物藿香蓟(Ageratum conyzoides L.)叶片中的镉含量最大值为77.01 mg/ kg[10] ;稻槎菜(Lapsana apogonoides) 在土壤镉浓度为2.04~2.89 mg/ kg 时,其地上部镉积累量为1.10 ~ 1.13mg/ m2[11] 。 以上研究结果表明,菊科植物在修复镉污染土壤中具有较好的应用前景,但这些研究只关注了植物富集重金属的方向,忽略了修复过程中的植物观赏性。 因此,从修复能力与景观修复效果相结合角度出发,选择应用于园林绿化的景观植物作为研究对象,可使既满足土壤修复条件又能达到景观效果的植物修复技术有更广的应用空间与更高的应用价值。
黄秋英(Cosmos sulphureus)又名硫华菊,菊科秋英属,一年生草本植物,花期为30~60 d[12] ,具有美观、生物量大、易养护等优点,是园林绿化中的热点植物之一。 目前对黄秋英的研究主要表现在化合物提取和耐盐抗旱等方面[13-15] ,而有关重金属对黄秋英影响的研究相对较少,且仅限于黄秋英的重金属富集能力研究,还未见重金属污染环境下黄秋英耐受能力的报道。 因此,本试验从修复能力与景观效果相结合的角度出发,选择应用于园林绿化的景观植物黄秋英为对象,研究不同质量浓度镉胁迫下黄秋英的耐受能力及其对黄秋英生理、光合特性的影响,明确其不同质量浓度镉胁迫下的富集能力,以期为景观植物的镉胁迫修复技术提供理论依据及技术支撑。
1 材料与方法
1.1 试验材料
供试黄秋英种子购自甘肃酒泉市蓝翔园艺种苗有限责任公司,千粒重9.1 g。
1.2 试验设计
试验于2022 年6 月15 日开始在吉林农业大学林学与草学学院温室进行。 试验采用盆栽方式,花盆规格为口径14.5 cm、底径10.1 cm、高度11.0 cm,栽培基质比例为草炭∶园土∶沙土=1∶2∶2,每盆装栽培基质500 g(干土)。
根据国家土壤污染风险管控标准( GB15618—2018,GB 36600—2018)并结合预试验所得结果,将试验土壤Cd2+ 浓度设置为:0、0.3、3、30、60、120、180 mg/ kg,共7 个处理,分别标记为C1、C2、C3、C4、C5、C6、C7。 供试Cd2+ 采用分析纯CdCl22.5H2 O 配制所得,以溶液形式浇入土壤中,保持土壤湿度一致,钝化平衡一个月开始试验。 黄秋英种子播种在育苗盘上,待幼苗长出2~4 片真叶时,选择生长良好且长势相同的幼苗移栽到花盆中,每盆3 株,栽植深度(5 cm)一致,并采用人工浇灌方式,每两天补水一次保持土壤湿润,于栽后15 d 和30 d 进行各项指标测定,重复3 次。
1.3 测定指标及方法
1.3.1 生理生化指标测定 超氧化物歧化酶(SOD)活性测定采用NBT 光还原法,过氧化物酶(POD)活性测定采用愈创木酚法;可溶性蛋白含量测定采用考马斯亮蓝G-250 染色法;丙二醛(MDA)含量测定采用硫代巴比妥酸法[16] ;叶片光合色素含量采用丙酮-乙醇(等体积比)混合液浸提法测定,计算单位鲜重、叶片的叶绿素a、叶绿素b、类胡萝卜素含量及叶绿素a/ b[17] 。
1.3.2 光合参数测定 镉胁迫15 d 和30 d,于晴天9 ∶00—11 ∶00 分别选择3 株长势相近植株,用Li-6800 光合作用仪(Li-Cor,Inc,USA)测定成熟叶片的光合参数,包括净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)。 测定过程中进入叶室的气体流速控制在500 μmol/ s,相对湿度控制在60%左右,参比室CO2 浓度控制在400 μmol/ L 左右,叶室温度在28~30 ℃之间,光强设定为1 400 μmol/ (m2s)。
1.3.3 叶绿素荧光参数测定 镉胁迫15 d 和30d,叶片经30 min 暗适应后测得初始荧光(Fo)和最大荧光(Fm),在光适应状态下测得实际荧光产量(Fo′)、稳态荧光产量(F′) 和最大荧光产量(Fm′)等荧光参数。 根据所测定荧光参数按照公式分别计算光系统Ⅱ的最大光化学量子产量(Fv/ Fm)、光合电子传递速率(ETR)、光化学淬灭系数(qP)、非光化学猝灭系数(NPQ):
Fv/ Fm=(Fm-Fo) / Fm ;
qP =(Fm-Fm′) / (Fm-Fo′) ;
NPQ=(Fm-Fm′) / Fm′ 。
1.3.4 镉含量测定 镉胁迫试验结束后,黄秋英整株收获,用ddH2 O 清洗,洗净后用吸水纸吸净表面水分,将植株分为根、茎、叶三部分。 每部分用天平称取10 g 样品,剪碎后装入聚四氟乙烯消解管中,加入硝酸5 mL 浸泡过夜后加入高氯酸0.5 mL 进行消解,再用石墨消解仪于180 ℃下消解45 min,将温度提高到220 ℃继续消解至黄棕色烟雾完全出尽,随后加入过氧化氢2 mL,继续加热至白烟出尽、溶液透明,消解结束后自然冷却至室温,定容至25 mL,使用可见分光光度计(型号:722N)进行镉含量测定[18] 。 通过标准曲线分别计算黄秋英根、茎、叶中的镉(Cd)含量,最后计算出黄秋英的镉富集系数(BCF) 与转移系数(TF)[19]。
富集系数(BCF)= 植物地上部镉含量或地下部镉含量/ 土壤中镉含量 ;转运系数(TF)= 植株地上部(茎叶)镉含量/地下部(根)镉含量 。
1.4 数据处理与分析
使用Microsoft Excel 2019 整理数据和图表制作,使用SPSS 20.0 软件进行单因素方差分析和差异显著性检验。
2 结果与分析
2.1 镉胁迫对黄秋英叶片生理生化指标的影响
2.1.1 对可溶性蛋白含量的影响 由表1 可知,不同镉浓度处理对黄秋英叶片可溶性蛋白含量的影响不同。 胁迫15 d 时,随着镉浓度增加,其可溶性蛋白含量呈先升后降趋势,C2与C1相比升高87.7%(P <0.05),C4、C5、C6、C7 与C1 相比下降37.4%、62.6%、74.7%、61.6%(P<0.05),C5、C6、C7较C4 显著降低。 胁迫30 d 时,随着镉浓度增加,
可溶性蛋白含量整体呈先升后降趋势,与C1 相比C2 升高22.1%(P<0.05),C3、C4、C5、C6、C7 分别降低27.8%、75.8%、83.6%、75.2%、90.4% (P <0.05),与C3 相比C4、C5、C6、C7 显著降低。 说明Cd 浓度0.3 mg/ kg 对黄秋英叶片可溶性蛋白的形成有促进作用;Cd 浓度增到3 mg/ kg 时,对黄秋英叶片可溶性蛋白的影响表现为,胁迫初期促进,后期抑制,并且随着胁迫浓度增加,抑制作用也逐渐加强。
2.1.2 对MDA 含量的影响 由表1 可知,不同镉浓度处理对黄秋英叶片MDA 含量的影响不同。 胁迫15 d 时,随着Cd 浓度增加,叶片MDA含量呈下降趋势,C5 和C7 处理达到最低,与C1相比均降低30.0%(P<0.05),C2、C4、C6 较C1 显著降低,C3 与C1 差异不显著。 胁迫30 d 时,随着Cd 浓度增加,叶片MDA 含量呈先升高后下降再升高的趨势:C3 较C1 升高26.5%(P<0.05);随后C4、C5 降低,但与C1 差异不显著;C6 达到最低,较C1 显著下降35.3%;C7 随后升高,与C6 相比增加50.0%,但与C1 差异不显著。 可以看出,Cd 胁迫初期,黄秋英叶片MDA 含量表现为降低,且随着胁迫浓度增加其生成受到抑制;Cd 胁迫后期则表现为低浓度促进MDA 生成,随着Cd 浓度增加则逐渐受到抑制。
2.1.3 对抗氧化酶活性的影响 由表1 可知,不同镉浓度处理对黄秋英叶片抗氧化酶活性的影响不同。 胁迫15 d 时,随着Cd 浓度增加,过氧化物酶(POD)活性整体呈先下降后升高趋势,C5 时达到最低,与C1 相比降低40.5%(P<0.05),C7 时达到最高,与C1 相比升高165.3%(P <0.05),其他浓度处理与C1 差异不显著;超氧化物歧化酶(SOD)活性呈先升后降趋势,C7 时达到最低,与C1 相比降低21.0%(P <0.05),其他浓度处理与C1 差异不显著。 胁迫30 d 时,随着Cd 浓度增加,POD 活性整体呈先下降后升高再下降的趋势,C2 时最低,与C1 相比降低65.0%,C4 与C1 相比显著升高,C5 处理达到最高,与C1 相比升高349.3%;SOD 活性呈先升后降趋势,与C1 相比C4、C6、C7 处理分别下降15.8%、26.8%、24.3%(P<0.05),其他处理与C1 差异不显著。
2.2 镉胁迫对黄秋英叶片光合色素含量的影响
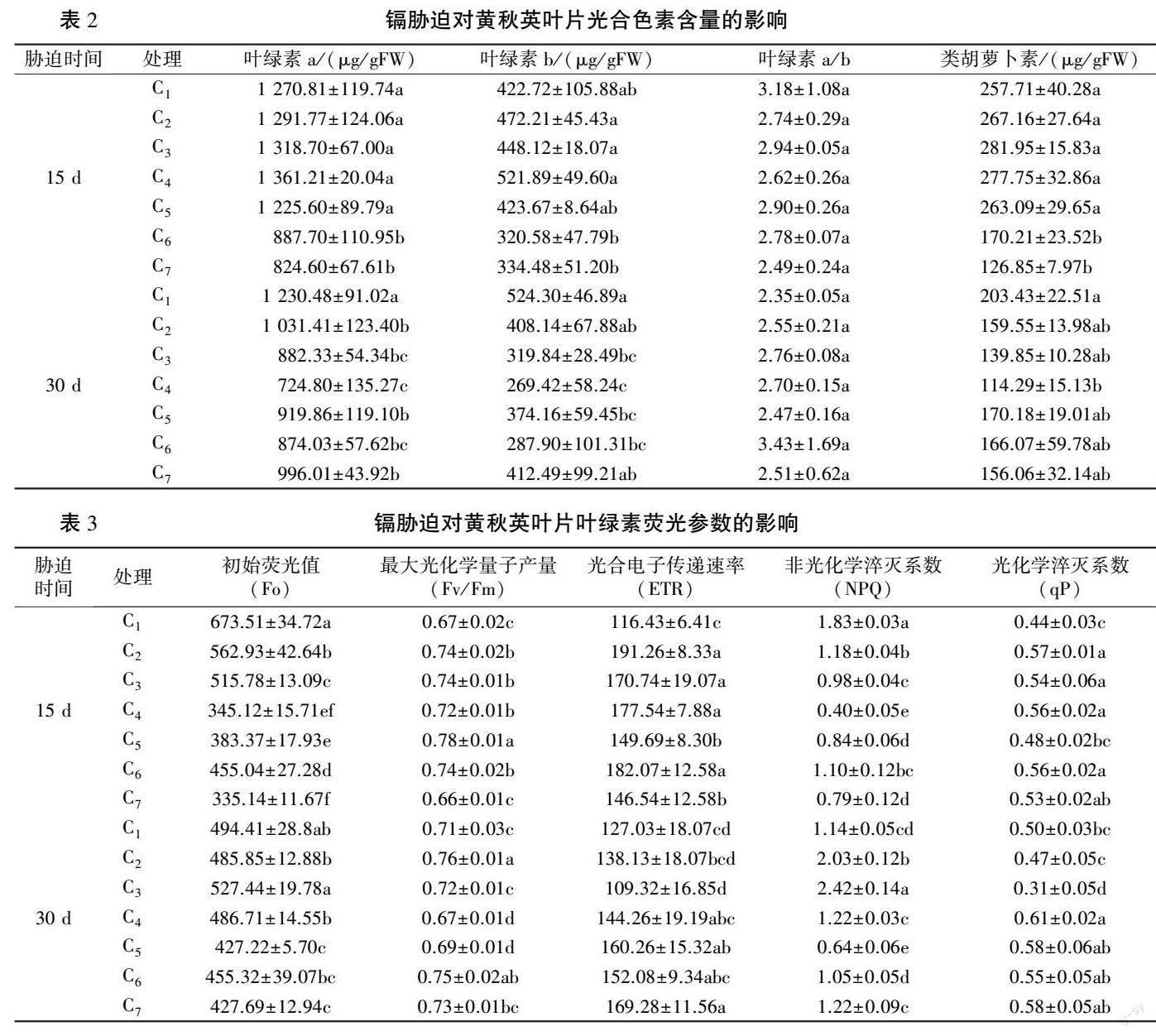
由表2 可知,不同Cd 浓度处理对黄秋英叶片光合色素含量的影响不同。 胁迫15 d 时,随着Cd 浓度增加,黄秋英叶片叶绿素a、叶绿素b、类胡萝卜素含量均呈先升后降趋势,三者中各增加处理与C1 差异均不显著,C6 较C1 分别下降30.1%(P<0.05)、24.1%、33.9%(P<0.05),且叶绿素b 含量C6 时达到最低,叶绿素a、类胡萝卜素含量C7 时达到最低, 比C1 分别降低35. 1%、50.8%(P<0.05)。
胁迫30 d 时,黄秋英叶片叶绿素a、葉绿素b、类胡萝卜素含量均呈现先下降后升高趋势,C4时达到最低,与C1 相比分别降低41.0%、48.6%、43.8%(P<0.05);与C1 相比,C2、C3、C5、C6、C7 叶绿素a 含量显著下降,类胡萝卜素含量差异不显著;C3、C5、C6 叶绿素b 含量与C1 相比显著降低,C2、C7 与C1 差异不显著。
胁迫15、30 d 时,各处理叶片叶绿素a/ b 与C1 差异均不显著。以上表明,胁迫15 d 时,黄秋英叶片叶绿素a、叶绿素b、类胡萝卜素含量,随着镉胁迫浓度增加,初期表现为促进,但效果未达到显著水平,Cd浓度增至120 mg/ kg 时表现为显著抑制;胁迫30d 时,随着镉胁迫浓度增加,抑制作用增强,Cd 浓度为30 mg/ kg 时达到最低,之后随着镉浓度增加抑制效果逐步减弱。
2.3 镉胁迫对黄秋英叶片叶绿素荧光特性的影响
由表3 可知,不同Cd 浓度处理对黄秋英叶片叶绿素荧光特性的影响不同。 镉胁迫15 d 时,随着Cd 浓度增加,初始荧光Fo 整体呈现逐步下降趋势,C7时降到最低,与C1相比降低50.2%,C5与C4相比略有上升但差异不显著,C6与C4相比显著升高达31.8%;最大光化学量子产量Fv/ Fm 呈现先升后降趋势,C5 时达到最高,与C1 相比显著升高16.4%,C7时达到最低,但与C1差异不显著;光合电子传递速率ETR 大体呈现先升高后下降趋势,C2时达到最高,与C1相比显著上升64.3%,C5和C7有所下降但仍显著高于C1;非光化学淬灭系数NPQ 大体呈现先下降后升高趋势,C4时降到最低,与C1相比显著降低78.2%,C6、C7与C4相比显著升高175.0%和97.5%,但与C1相比分别显著下降39.9%和56.8%;光化学淬灭系数qP 大体呈现先升高后下降再升高的变化趋势,C2时达到最高,
较C1 显著升高29.5%,C5 与C2 、C3 、C4 、C6 相比显著下降,但与C1和C7差异不显著。镉胁迫30 d 时,随着Cd 浓度增加,Fo 呈现先升高后下降趋势,C3 时达到最高,但与C1 差异不显著,C5时最低,与C1相比显著降低13.6%,与C6和C7差异不显著;Fv/ Fm 呈现先升后降再升再降的变化趋势,C2 时达到最高,与C1 相比显著升高7.1%,C4时最低,与C1相比显著降低5.6%,C6与C1相比升高显著,C7 与C1 差异不显著;ETR 大体呈现先升高后下降再升高的变化趋势,C3 时最低,但与C1差异不显著,C7时达到最高,与C1相比显著提升33.3%;NPQ 整体呈现先升高后下降趋势,C3时达到最高,与C1相比显著升高112.3%,C5时最低,与C1相比显著降低43.9%,C6、C7略有回升但与C1 差异不显著;qP 呈现先下降再升高趋势,C3 时降到最低,与C1 差异显著,C4 时达到最高,与C1相比显著上升22.0%,C5、C6、C7与C1、C4差异不显著。
2.4 镉胁迫对黄秋英叶片光合特性的影响
由图1 可知,不同浓度镉处理对黄秋英叶片光合参数的影响不同。 胁迫15 d 时,随着Cd 浓度增加,黄秋英叶片的蒸腾速率(Tr)、净光合速率(Pn)、气孔导度(Gs)大体呈现先升高后下降趋势,胞间CO2 浓度(Ci)无显著差异。 其中,Tr 在C2 时达到最高,与C1 相比显著上升68.7%,C7 时降到最低,与C1 差异不显著;Pn 在C3 时达到最高,与C1 相比显著上升91.1%,C7 时降到最低,与C1 差异不显著;Gs 在C3 时达到最高,与C1 相比显著上升113.0%,C7 与C1 差异不显著。
胁迫30 d 时,随着Cd 浓度增加,黄秋英叶片的Tr、Pn、Gs 大体呈现先下降再升高趋势,Ci 呈现先升高后下降再升高的变化趋势。 其中,Tr 在C3 时降到最低,与C1 相比显著下降29.3%,C7 时达到最高,与C1 相比显著升高25.3%;Pn 在C3时降到最低,与C1 相比显著下降32.7%,C5 时达到最高,与C1 相比显著升高18.2%ê.Gs 在C3 时降到最低,与C1 相比显著降低35.7%,C5 时达到最高,与C1 相比升高28.6%;Ci 在C6 时降到最低,与C1 相比显著下降9.2%,C7 时达到最高,但与C1 差异不显著。
2.5 镉胁迫对黄秋英镉含量与镉积累的影响
由表4 可知,随着镉胁迫浓度增加,黄秋英根、茎、叶及地上部镉含量均呈现上升趋势,C5、C6处理各器官镉含量分布顺序为茎>根>地上部>叶,,C1、C2、C3、C4 为茎>地上部>根>叶,C7 为根>茎>地上部>叶。 C2 ~ C7 这6 个处理黄秋英根系镉含量分别是C1 的2.50、22.39、185.56、456.50、699.61 倍和854.61 倍,C4 ~C7 均与C1 差异显著;地上部镉含量分别是C1 的3.05、24.95、244.37、380.58、548.05 倍和611.47 倍,C4 ~C7 均与C1 差异显著。 镉浓度为0.3、3、30、60 mg/ kg(C2、C3、C4、C5)时,其地上部富集系数大于1,但镉含量均未达到镉超富集植物标准(100 mg/ kg),120、180mg/ kg(C6、C7)时,黄秋英根、茎及地上部镉含量均达到镉超富集植物标准,但富集系数与转运系数均小于1,因此判定黄秋英是镉富集植物。
由表5 可知,随着镉浓度增加,其他各处理黄秋英地上部与地下部镉积累量均高于C1,其中地下部镉积累量呈现上升趋势,地上部镉积累量呈现先升后降趋势。 C2 和C3 地下部镉积累量分别是C1 的2.35 倍和17.40 倍,但差异不显著;C4、C5、C6、C7 分别是C1 的182.38、314.56、346.82、607.85 倍,差异达到显著水平;C2 ~ C7 这6 个处理地上部镉积累量分别是C1 的1.30、17.13、127.35、172.68、295.52、68.31 倍。 地下部镉积累量在C7 ( 180 mg/ kg) 时达到最高, 为151. 96μg/ 株,地上部鎘积累量在C6(120 mg/ kg)时达到最高,为14.78 μg/ 株。
3 讨论与结论
3.1 镉胁迫对黄秋英叶片生理生化指标的影响
重金属镉(Cd)具有很强的生物毒性,Cd2+ 在植物体内积累会导致植物细胞氧化应激,并积累大量的活性氧(ROS),引起膜脂过氧化,细胞膜透性增强,破坏膜结构,致使植物自身的生理生化系统紊乱[20] 。 而有关研究表明,植物体会通过自身抗氧化系统去消除活性氧以维持正常的生理代谢与稳定的内环境[21] 。 MDA 是膜脂过氧化的重要产物之一,膜脂过氧化的程度越深,MDA 含量就越高[22] 。 本试验中,胁迫15 d 时,随着Cd 浓度增加,黄秋英叶片MDA 含量逐渐下降;胁迫30 d时MDA 含量在C3 时达到最高,后随着胁迫浓度增加逐渐降低,C7 下MDA 含量与C1 相比差异不显著。 表明高浓度和长时间的Cd 胁迫对黄秋英叶片的细胞膜破坏不严重,这与陈一博[23] 的镉胁迫在早开堇菜上的研究结果不一致,推测是由不同植物对镉的耐受能力不同所致。
POD 与SOD 是植物体清除ROS 的主要抗氧化酶。 本试验中,镉胁迫15 d 时,黄秋英叶片POD 活性整体呈先下降后升高趋势,SOD 呈现先升高后下降趋势。 其中,POD 活性在C5时降到最低,此时SOD 活性与C1相比略有提升;POD 活性在C7时达到最高,此时SOD 活性降到最低。 胁迫30 d 时,POD 活性呈现先下降后升高再下降的变化趋势,SOD 活性呈现先升高后下降趋势。 其中,POD 活性在C2时降到最低,此时SOD 与C1相比略有提升;C4下SOD 活性较C1 显著降低,此时POD 活性与C1相比显著提高;之后随着Cd 浓度增加,POD 和SOD 活性均呈降低趋势,C6、C7 下POD 活性仍高于C1,而SOD 活性则显著降低。表明黄秋英在Cd 胁迫下以提高POD 活性作为清除体内ROS 的主要手段,这与林立金等[24] 的试验结果一致,而SOD 则作为辅助调节。 在高浓度和长时间Cd 胁迫下,随着毒害的加深,POD 活性仍高于C1,由此可见黄秋英对于Cd 胁迫有较强的耐受性,能够在土壤Cd 污染环境下维持自身内环境的平衡。
可溶性蛋白是植物体内重要的渗透调节物质,在逆境条件下可溶性蛋白的积累对维持植物细胞正常的生理活动有着重要意义[25] 。 本试验中,胁迫15 d 与30 d 时,随着Cd 浓度增加,黄秋英叶片可溶性蛋白含量先升高后下降,浓度为C2时达到最高,之后逐渐降低。 这表明低浓度Cd胁迫下,黄秋英能够通过提高可溶性蛋白含量来抵御Cd 的毒害,但随着胁迫浓度增加,可溶性蛋白合成受到抑制,含量减少。 该结果与刘翰升等[18] 对万寿菊的试验结果一致,但同时期POD与SOD 的作用能够很好地降低黄秋英体内的ROS 含量,降低Cd2+ 对细胞膜的毒害,保证黄秋英膜系统不遭到严重破坏。
3.2 镉胁迫对黄秋英叶片光合色素含量和光合特性的影响
光合作用是植物维持生命活动的基础,长期暴露在Cd 环境中的植物,其体内光合系统会遭到破坏,导致光合能力降低,从而对植物正常的生命活动造成影响[26] 。 本试验结果表明,短期(15d)胁迫下低浓度Cd 能够促进黄秋英叶片的光合作用,而随着Cd 浓度增加,毒害随之加深,促进逐渐转为抑制。 其中,Tr、Gs 和叶绿素a、叶绿素a/ b、类胡萝卜素均在Cd 浓度为180 mg/ kg 时其值降到最低,叶绿素b、Ci 有所下降,表明高浓度Cd 胁迫导致黄秋英叶片Tr、Gs 下降的同时减少光合色素的合成,导致光合速率(Pn)降低。 短期高浓度Cd 胁迫虽然能够抑制黄秋英的光合速率,但这种抑制不显著,光合作用仍能正常运行;而长期(30 d)Cd 胁迫下,Tr、Pn、Gs 均在Cd 浓度为3 mg/ kg 时降到最低,叶绿素a、叶绿素b 含量显著降低,之后随着胁迫浓度增加呈现升高趋势。这与王菲等[27] 的试验结果不一致。 推测其原因可能是,此浓度下的黄秋英叶片抗氧化酶系统调节速度缓慢,ROS 没有得到及时消除,导致黄秋英叶片细胞膜系统遭到破坏,同时影响了光合色素合成,进而造成叶片光合速率降低,后随着胁迫浓度增加,黄秋英抗氧化酶系统调节速度加快,细胞膜系统恢复正常,叶片光合速率也与C1 差异不显著。
3.3 镉胁迫对黄秋英叶片叶绿素荧光特性的影响
初始荧光Fo 上升可能反映植物PSⅡ受到不可逆的破坏或可逆性损伤;Fv/ Fm 是指PSⅡ最大光化学量子产量,即PSⅡ反应中心内禀光能转换效率,它能够反映PSⅡ的受损程度[28] 。 本试验中,Cd 胁迫15 d 时,随着Cd 浓度增加,Fo 逐渐降低;Cd 浓度在180 mg/ kg 时Fv/ Fm 降到最低,但与C1差异不显著。 Cd 胁迫30 d 时,Fo 在镉浓度3 mg/ kg 时达到最高,后随着胁迫浓度增加而降低;Fv/ Fm 在镉浓度30 mg/ kg 时降到最低,后随着胁迫浓度增加而升高。 这表明短期(15 d)低浓度Cd 胁迫对黄秋英PSⅡ会产生有利影响,而高浓度胁迫对其为不利影响,但不显著;而长期(30d)中低浓度Cd 胁迫会对黄秋英PSⅡ造成损害。该结果与张敏等[29] 的试验结论不一致,其原因可能是由此时的抗氧化酶系统调节速度缓慢而造成,后随着胁迫浓度增加其响应速度加快,Cd2+ 对PSⅡ的损害得到抑制与修复。
非光化学淬灭系数NPQ 反映的是PSⅡ天线色素吸收的光能不能用于光合电子传递而以热的形式耗散掉的光能部分;光化学淬灭系数qP 指PSⅡ天线色素吸收光能后用于光合作用电子传递的份额,反映PSⅡ反应中心的开放程度;ETR能够反映植物体光反应中心PSⅡ在光适应条件下真实的电子传递速率[30] 。 本试验中,Cd 胁迫15 d 时,NPQ 随Cd 浓度增加先降低后升高,但回升后仍低于C1;qP 则在Cd 浓度为60 mg/ kg 时与C1差异不显著,其他浓度下均显著高于C1;ETR在Cd 浓度最高(180 mg/ kg) 时仍显著高于C1。这表明短期Cd 胁迫黄秋英主要是通过提高光合电子传递速率和降低以热能形式的耗散来抵御Cd2+对其光系统的毒害。 Cd 胁迫30 d 时,NPQ在Cd 浓度3 mg/ kg 时升至最高,后随着胁迫浓度增加而降低,Cd 浓度120 mg/ kg 时又出现上升趋势,180 mg/ kg 时高于C1 但差异不显著;ETR 在Cd 浓度3 mg/ kg 时低于C1,其余均高于C1,且高浓度胁迫时上升趋势增加;qP 在Cd 浓度3 mg/ kg时降至最低,后呈升高趋势且均高于C1。 此结果与张栋栋等[31] 的研究相近。 贾中民等[32] 在秋华柳(Salix variegata)上的试验中指出,镉会使其光合机构受到毒害,PSⅡ原初醌受体的光化学还原效率被Cd 抑制,降低PSⅡ向PSⅠ的电子传递,使得叶片吸收的光能在到达反应中心之前,主要以热损耗的形式散失,从而增强植物自身的光保护能力,避免或减轻了光抑制甚至光氧化对植物自身光合机构的损害,使植物能够维持一定的光合作用水平。 本试验结果表明,长时间低浓度Cd胁迫限制了黄秋英的电子传递速率,导致光反应速率降低,反应中心的开放程度减小,天线色素吸收的光能以热能形式散发的比例增加,此时光反应中心遭到损害,而后随着抗氧化酶系统调节能力的提升,光合机构受到的毒害降低,这种现象得到缓解;长期高浓度Cd 胁迫虽然能够对黄秋英PSⅡ造成影响,但通过自身的调节机制,使得其能够维持一定的光合作用水平。
3.4 镉胁迫对黄秋英镉含量与镉积累的影响
目前国内关于镉超富集植物的鉴定标准为地上部镉浓度达到临界值100 mg/ kg 且转运系数大于1[33] 。 本试验结果表明,黄秋英各器官镉含量在镉处理浓度<180 mg/ kg 时分布顺序为茎>根>叶;镉处理浓度≥180 mg/ kg 时为根>茎>叶。 黄秋英根、茎、叶及地上部镉含量均随着胁迫浓度增加而升高,当浓度≥120 mg/ kg 时黄秋英根、茎及地上部镉含量均超过镉超富集植物的临界值(100 mg/ kg),但此时转运系数小于1,这与林立金[24] 等的试验结果一致。 黄秋英地下部与地上部镉积累量随着胁迫浓度增加分别呈现逐渐升高和先升后降趋势,地下部镉积累量在Cd 浓度180mg/ kg 时达到最大值(151.96 μg/ 株),地上部镉积累量在Cd 浓度120 mg/ kg 时达到最大值(14.78 μg/ 株)。 由此看出,黄秋英为镉积累植物,高浓度下仍有较好的富集效果,对镉污染土壤的治理与修复有着重要作用。
综上所述,黄秋英在高浓度镉胁迫下具有较高的耐受力,同时具备较强的镉富集能力,在镉污染环境中能够维持正常的生命活动,可以作为一种具有景观修复性的植物材料用于Cd 污染土壤修复。
参 考 文 献:
[1] 宋伟,陈百明,刘琳. 中国耕地土壤重金属污染概况[J]. 水土保持研究,2013,20(2):293-298.
[2] 中华人民共和国环境保护部,中华人民共和国国土资源部.全国土壤污染状况调查公报[J]. 中国环保产业,2014(5):10-11.
[3] He B,Yun Z J,Shi J B,et al. Research progress of heavymetal pollution in China: sources,analytical methods,status,and toxicity[J]. Chinese Science Bulletin,2013,58(2):134-140.
[4] 赵娜. 吉林省稻田土壤生境恶化问题的研究[D]. 长春:吉林农业大学,2015.
[5] 赵升,欧阳凯,罗绪强,等. 贵阳市高雁垃圾填埋场周边土壤和农作物重金属污染特征及健康风险评价[J]. 山东农业科学,2023,55(2):92-99.
[6] 文晓阳,饶巍,李春萍,等. 铅胁迫对不同基因型烟草铅吸收、运转及积累的影响[J]. 山东农业科学,2022,54(9):106-112.
[7] 刘少文,焦如珍,董玉红,等. 土壤重金属污染的生物修复研究进展[J]. 林业科学, 2017,53(5):146-155.
[8] 赵晓峰,雷梅,陈同斌. 中国镉超富集植物的物种、生境特征和筛选建议[J]. 环境科学,2023,44(5):2786-2798.
[9] 张云霞,周浪,肖乃川,等. 鬼针草(Bidens pilosa L.)对镉污染农田的修复潜力[J]. 生态学报,2020,40(16):5805-5813.
[10] 张云霞,宋波,宾娟,等. 超富集植物藿香蓟(Ageratum cony ̄zoides L.)对镉污染农田的修复潜力[J]. 环境科学,2019,40(5):2453-2459.
[11] 林立金,石軍,刘春阳,等. 稻田冬季杂草稻槎菜的镉积累特性研究[J]. 华北农学报, 2016,31(2):146-152.
[12] 陈林. 园林花卉[M]. 重庆:重庆大学出版社,2015:321.
[13] Iwashina T, Amamiya K, Kamo T, et al. 2′ ̄Hydroxylated 3 ̄Deoxyanthocyanin from the flowers of Cosmos sulphureus culti ̄vars[ J]. Natural Product Communications, 2019, 14 ( 9):1934578×1987621.
[14] 崔京荣,刘桂林. Na2 CO3 胁迫对5 种草本花卉种子萌发的影响[J]. 种子,2018,37(2):39-42.
[15] 吴晓凤,倪沛,杨涛,等. 10 种菊科植物的抗旱性与抗盐性评价[J]. 生态学杂志,2018,37(7):1959-1968.
[16] 国伟强,杜雪,彭玉兰,等. Cd、Pb 复合胁迫对狼尾草种子萌发及幼苗生长的影响[J]. 种子,2021,40(11):91-96.
[17] 朱景乐,杜红岩,李芳东,等. 杜仲叶片光合色素含量测定方法筛选[J]. 河南农业大学学报,2013,47(5):557-561.
[18] 刘翰升,赵春莉,刘玥,等. Cd 胁迫对波斯菊种子萌发、幼苗耐性及富集的影响[J]. 河南农業科学,2020,49(5):126-133.
[19] 周强英,黄泽梅,陈瑶. 铅镉复合胁迫下黄葛树和女贞的生长生理及吸收特性研究[J]. 西南林业大学学报(自然科学),2019,39(6):33-40.
[20] 赵晶晶,詹万龙,周浓. 非生物胁迫下植物体内活性氧和丙酮醛代谢的研究进展[J]. 南方农业学报,2022,53(8):2099-2113.
[21] 朱秀红,程红梅,季柳洋,等. 锌、镉及其复合胁迫下白花泡桐幼苗的生理及富集特征[J]. 生物工程学报,2021,37(7):2463-2473.
[22] Monteiro M S, Santos C, Soares A, et al. Assessment of bio ̄markers of cadmium stress in lettuce[ J]. Ecotoxicology andEnvironmental Safety,2008,72(3):811-818.
[23] 陈一博. 铅、镉单一及复合胁迫对早开堇菜生理及富集特性的影响[D]. 哈尔滨:东北林业大学,2022.
[24] 林立金,马倩倩,石军,等. 花卉植物硫华菊的镉积累特性研究[J]. 水土保持学报, 2016,30(3):141-146.
[25] 向亮,王艳杰,陈佳勃,等. 盐和重金属复合胁迫对翅碱蓬萌发与生长的影响及调控措施[J]. 生态学报,2023,43(8):3307-3318.
[26] 熊敏先,吴迪,许向宁,等. 土壤重金属镉对高等植物的毒性效应研究进展[J]. 生态毒理学报,2021,16(6):133-149.
[27] 王菲,肖雨,程小毛,等. 镉胁迫对吊兰及银边吊兰生长及镉富集特性的影响[J]. 应用生态学报,2021,32(5):1835-1844.
[28] 张守仁. 叶绿素荧光动力学参数的意义及讨论[J]. 植物学通报,1999,16(4):444-448.
[29] 张敏,李燕,宁朋,等. 镉胁迫对八角金盘生长及生理特性的影响[J]. 分子植物育种, 2023,21(3):978-987.
[30] 钱永强,周晓星,韩蕾,等. Cd2+ 胁迫对银芽柳PSⅡ叶绿素荧光光响应曲线的影响[J]. 生态学报,2011,31(20):6134-6142.
[31] 张栋栋,李萌,甘龙,等. 重金属Cd-Cu 复合污染对苍耳生长及叶绿素荧光特性的影响[J]. 武汉大学学报(理学版),2019,65(1):66-76.
[32] 贾中民,程华,魏虹,等. 三峡库区岸生植物秋华柳对镉胁迫的光合响应[J]. 林业科学,2012,48(6):152-158.
[33] 汪京超. 基于转录组学的油菜镉胁迫响应机制研究[D].重庆:西南大学,2016.
