干旱低温复合胁迫下黄腐酸钾对西葫芦幼苗生长和生理生化特性的影响
2024-04-01孔繁荣李海平郭文忠武月胜郭建林周可杰
孔繁荣 李海平 郭文忠 武月胜 郭建林 周可杰


摘要:本试验以西葫芦‘农园1 号为材料,设置7 个处理,分别是对照(CK,无干旱低温胁迫且不浇灌黄腐酸钾)、DL(干旱低温胁迫下不浇灌黄腐酸钾)和T1、T2、T3、T4、T5(干旱低温胁迫下黄腐酸钾浇灌浓度分别为0.020%、0.035%、0.05%、0.065%、0.08%),研究干旱低温胁迫下黄腐酸钾对西葫芦幼苗生长和生理生化特性的影响。 结果表明:干旱低温胁迫下西葫芦幼苗的干鲜重、脯氨酸含量和SOD、POD、CAT 活性以及光合能力均受到抑制,不同浓度黄腐酸钾处理均可缓解干旱低温对西葫芦幼苗生长和生理生化指标的抑制作用。 其中T3 处理效果最佳,其幼苗叶面积、全株鲜重、全株干重、脯氨酸含量和SOD、POD、CAT 活性与DL 相比分别显著增加13.89%、16.20%、17.43%、19.43%和13.48%、7.13%、16.28%,MDA 含量降低14.37%,叶绿素总量、净光合速率显著增加19.66%和17.95%,叶片的Fv/ Fm、ΦPSⅡ、ETR、qP 显著升高4.25%、17.57%、14.85%和14.01%,Fo、NPQ 分别降低5.30%、15.79%,即减轻了干旱低温胁迫对叶片光合器官的損伤。 综上看出,黄腐酸钾可以在一定程度上缓解干旱低温胁迫对西葫芦幼苗生长和生理生化指标的抑制作用,以0.05%黄腐酸钾处理(T3)效果较为显著。
关键词:西葫芦;黄腐酸钾;干旱;低温;幼苗生理
中图分类号:S642.6文献标识号:A文章编号:1001-4942(2024)01-0074-07
西葫芦(Cucurbita pepo L.) 在我国栽培普遍,节能日光温室的引进和发展更使冬春季日光温室西葫芦栽培面积迅速增加[1] 。 尽管日光温室西葫芦种植面积不断增加,但在水、肥和光照管理等方面仍按照露地模式进行,导致病害发生严重、品质变差、产量下降、经济效益不佳[1] 。 北方设施栽培中,低温也会导致作物生长不良,降低作物产量[2]。
应用外源物质缓解或者减轻逆境给作物带来的伤害已有研究[3] :盐胁迫下,水杨酸溶液处理的种子发芽率提高,对根和茎的抑制效应减轻,提高黄瓜的耐盐性[4] ;喷施海藻酸钠寡糖和赤霉素复配剂,可快速恢复亚低温逆境下的番茄生长,促进养分吸收和光合作用,增加有机物的积累量,提高果实产量和品质[5] ;喷施适量浓度的脱落酸可以减轻铅胁迫对植株的毒害作用,增强其对铅胁迫的抵御能力,增强植物的光合作用[6] ;喷施甜菜碱可以提高干旱胁迫下茄子幼苗的生长,降低MDA 含量,提高茄子的耐旱性[7] ;外源褪黑素通过提高清除活性氧的能力和增强光合作用,缓解低温胁迫对茄子幼苗的伤害[8] 。
黄腐酸(fulvic acid,FA) 具有分子量小、容易被植物吸收利用、生理活性强、易溶于水等特点[9] 。 刘玉丰等[10] 研究发现,黄腐酸钾可以增加盐碱胁迫下甜瓜幼苗的生物量、净光合速率,缓解盐碱胁迫对甜瓜幼苗的抑制作用。 刘彩娟等[11]研究表明,黄腐酸显著增加干旱胁迫下温室黄瓜的叶绿素含量、净光合速率,促进干物质积累,提高黄瓜的抗旱性。 张彩凤等[12] 研究发现,低温胁迫下黄腐酸钾能够增强植物的抗氧化酶活性,抑制MDA 含量升高,提高净光合速率,增强植物的抗寒性。 可以看出,外源物质在作物抗逆性方面已有较多研究,但在干旱低温共同胁迫下外源物质对作物生长和生理影响的研究还鲜见报道。 为此,本试验以西葫芦为试材,研究不同浓度黄腐酸钾对干旱低温胁迫下西葫芦幼苗生长和生理生化指标的影响,以期为生产中提高西葫芦幼苗非生物抗性提供理论依据。
1 材料与方法
1.1 试验材料
供试西葫芦品种为‘农园1 号。 黄腐酸钾由山西林海腐植酸科技有限公司提供,腐植酸含量≥50%、黄腐酸≥47%、K2O≥12%。
1.2 试验设计
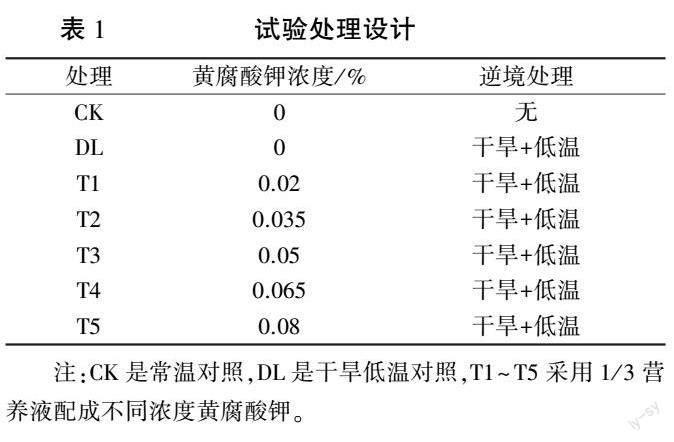
试验于2021 年9—10 月在山西省晋中市东阳镇(37°32′N,112°40′E) 日光温室(昼温23 ~28 ℃,夜温13 ~16 ℃,自然光照)中进行。 将发芽一致的种子播种于草炭∶珍珠岩(V/ V)= 7∶2 的21 孔穴盘中,待幼苗长至一叶一心时,将长势一致的幼苗分为7 组,每组21 株。 试验设置7 个处理,见表1。 T1—T5 处理每天浇灌相应浓度的黄腐酸钾溶液,连续浇灌5 天,之后干旱(田间持水量的60%±5%)胁迫5 天,再低温胁迫5 天。 低温结束后缓苗3 天取样测定相关指标。干旱处理:用称重法控制水分,参考李建明等[13] 的方法进行。 通过称重记录幼苗的蒸腾蒸发量,对照(CK)按照处理前一天100%蒸发量补充水分,DL、T1~T5 处理按照60%±5%蒸发量补充水分。
低温处理:参考董绪兵等[14] 的方法在人工气候室进行。 其条件为:昼温(7±1) ℃,12 h;夜温(5±1) ℃,12 h,光照400 μmol/ (m2s)。
1.3 测定指标及方法
1.3.1 生长指标测定 用直尺测量株高(茎基部到生长点的距离)、叶长、叶宽;游标卡尺测量茎粗(基部粗度);电子天平(0.01 g)称量西葫芦幼苗全株鲜重,然后置于烘箱105 ℃ 杀青30 min、75 ℃烘干至恒重称干重。 叶面积计算公式[15]为:S=0.578 1+0.774 8LB。 式中,S 是单片叶叶面积,L 是叶长,B 是最大叶宽。
1.3.2 生理指标测定 参考高俊凤等[16] 的方法测定。 其中,乙醇提取法测叶绿素含量,酸性茚三酮比色法、蒽酮比色法和考马斯亮蓝法分别测定脯氨酸、可溶性糖和可溶性蛋白含量;采用氮蓝四唑还原法、愈创木酚比色法、紫外吸收法、硫代巴比妥酸比色法分别测定SOD、POD、CAT 活性和MDA 含量。
1.3.3 光合指标测定 于9∶00—11∶00 用LI-6800 光合仪在自然光下测量各处理幼苗自上向下数第二片叶的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间二氧化碳浓度(Ci),测定时设置光量子通量密度为500 μmolm-2s-1,湿度为50%。
1.3.4 叶片荧光指标测定 用LI-6800 光合仪荧光叶室测定光适应下每株幼苗自上向下第二片叶的荧光参数:PSⅡ实际光化学效率(ΦPSⅡ)、电子传递效率(ETR)、光化学猝灭系数(qP)和非光化学猝灭系数(NPQ)等;暗适应1 h 后,用LI-6800 光合仪荧光叶室测定暗适应下荧光参数:初始荧光(Fo)、PSⅡ最大光化学量子产量(Fv/ Fm)。
1.4 数据处理与分析
用Microsoft Excel 2016 进行数据处理,SPSS25.0 软件进行数据分析。
2 结果与分析
2.1 干旱低温胁迫下黄腐酸钾对西葫芦幼苗生物量的影响
由表2 可见,干旱低温胁迫(DL)下西葫芦幼苗株高、茎粗、叶面积、全株鲜重和全株干重与对照(CK) 相比分别降低12.56%、12.07%、25.97%、20.00%、25.57%(P<0.05),说明干旱低温胁迫抑制西葫芦幼苗生长。 干旱低温胁迫下不同浓度黄腐酸钾处理西葫芦幼苗株高、茎粗、叶面积、全株鲜重和全株干重与DL 相比均有不同程度的提高,且随黄腐酸钾浓度升高呈现先升高再降低趋势,其中T3 处理增幅最大,与DL 相比分别增加9.77%、10.46%、13.89%、16.20%和17.43%。 表明黄腐酸钾可以一定程度上减轻干旱低温胁迫对西葫芦幼苗生长的抑制,浓度0.05%处理(T3)的效果最好。
2.2 干旱低温胁迫下黄腐酸钾对西葫芦幼苗渗透调节物质的影响
由表3 可知,对照(CK)相比,干旱低温胁迫(DL)下幼苗叶片的脯氨酸、可溶性糖和可溶性蛋白含量分别提高10.61%、20.67%、14.63%,且后两者达显著水平。 干旱低温胁迫下不同浓度黄腐酸钾处理西葫芦幼苗叶片的脯氨酸、可溶性糖和可溶性蛋白含量较DL 处理均有不同幅度的提高,并且随黄腐酸钾浓度增加表现为先升高再降低趋势,其中T3 处理的脯氨酸、可溶性糖、可溶性蛋白含量最高,与DL 相比分别提高19. 43%、19.02%、15.95%(P <0.05)。 说明黄腐酸钾在一定程度上增加干旱低温胁迫下西葫芦幼苗叶片的脯氨酸、可溶性糖和可溶性蛋白积累,浓度0.05%处理(T3)的效果最显著。
2.3 干旱低温胁迫下黄腐酸钾对西葫芦幼苗丙二醛含量和抗氧化酶活性的影响
由表4 可见,干旱低温胁迫处理(DL)西葫芦幼苗叶片的丙二醛(MDA)含量与对照(CK)相比显著升高,说明干旱和低温条件对西葫芦造成伤害,膜脂过氧化产物积累。 干旱低温胁迫下不同浓度黄腐酸钾处理西葫芦幼苗叶片的MDA 含量与DL 相比均有不同程度的降低,并随黄腐酸钾浓度增加表现为先降低再升高趋势,其中T3 处理的MDA 含量最低,比DL 降低14.37%(P<0.05)。表明黄腐酸钾可以降低叶片MDA 含量,减轻干旱低温胁迫对西葫芦幼苗的伤害,最适浓度为0.05%(T3)。
由表4 可见,干旱低温胁迫(DL)下西葫芦幼苗叶片的SOD、POD、CAT 活性显著高于对照(CK),分别高11.79%、9.89%和5.79%。 干旱低温胁迫下不同浓度黄腐酸钾处理西葫芦幼苗叶片的SOD、POD、CAT 活性均不同程度高于DL 处理,其中浓度0.05%处理最高,分别比DL 处理增加13.48%、7.13%、16.28%。 表明黄腐酸钾可以提高西葫芦幼苗的抗氧化酶活性,缓解干旱低温胁迫造成的伤害。
2.4 干旱低溫胁迫下黄腐酸钾对西葫芦幼苗叶片光合色素含量的影响
由表5 可知,与对照(CK)相比,干旱低温胁迫(DL)下西葫芦幼苗叶片的叶绿素a、叶绿素b、类胡萝卜素含量和叶绿素总量分别降低28.35%、25.72%、20.00%、27.18%(P<0.05)。 干旱低温胁迫下不同浓度黄腐酸钾处理西葫芦幼苗叶片的叶绿素a、叶绿素b、叶绿素总量和类胡萝卜素含量均比DL 处理有不同程度的提高,且随黄腐酸钾浓度增加表现为先增加再降低趋势,以T3 处理的叶片光合色素含量增幅最大,比DL 处理分别增加18.69%、19.23%、19.66%(P<0.05)和12.50%。表明黄腐酸钾可有效增加干旱低温胁迫下西葫芦幼苗叶片的光合色素含量,最适浓度为0.05%(T3)。
2.5 干旱低温胁迫下黄腐酸钾对西葫芦幼苗叶片光合指标的影响
由表6 可知,干旱低温胁迫处理(DL)西葫芦幼苗叶片的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间二氧化碳浓度(Ci)均显著低于对照(CK),分别降低19.06%、18.36%、17.95%和14.35%。 干旱低温胁迫下不同浓度黄腐酸钾处理西葫芦幼苗叶片的Pn、Tr、Gs、Ci 与DL 处理相比均有不同程度的提高,并随黄腐酸钾浓度增加呈先升后降趋势,其中T3 处理的各指标值均最高, 分别比DL 处理提高17. 95%、12.41%、12.50%、7.37%(P<0.05)。 表明黄腐酸钾一定程度上能够缓解干旱低温胁迫对西葫芦幼苗光合的抑制,浓度0.05%处理(T3)缓解作用最显著。
2.6 干旱低温胁迫下黄腐酸钾对西葫芦幼苗叶片荧光参数的影响
由表7 可知,干旱低温胁迫(DL)下西葫芦幼苗叶片的Fv/ Fm、ΦPSⅡ、ETR、qP 显著低于对照(CK), 分别降低4. 37%、18. 69%、20. 38% 和15.29%,Fo、NPQ 分别显著升高8.02%、19.80%,表明干旱低温胁迫造成叶片PSⅡ部分失活或伤害,光反应能力降低,光合效率、光能利用效率降低。 干旱低温胁迫下不同浓度黄腐酸钾处理均能不同程度缓解西葫芦幼苗叶片Fv/ Fm、ΦPSⅡ、ETR、qP 的降低和Fo、NPQ 的升高。 总体上来看,叶片Fv/ Fm、ΦPSⅡ、ETR、qP 随黄腐酸钾浓度增加均表现为先升高再降低趋势,Fo、NPQ 则是先降低再升高趋势。 其中,与DL 相比,T3 处理(0.05%)西葫芦幼苗叶片的Fv/ Fm、ΦPSⅡ、ETR、qP 最大,分别增加4. 25%、17. 57%、14. 85% 和14.01%,Fo、NPQ 分别降低5.30%和15.79%。 说明适宜浓度的黄腐酸钾可以一定程度上降低干旱低温胁迫对西葫芦幼苗PSⅡ的伤害,提高光合效率。
3 讨论
逆境胁迫下植物的株高、茎粗、干鲜重受到抑制,生长发育受阻[17] 。 本研究表明,干旱低温胁迫下西葫芦幼苗生长发育受到抑制,且随着黄腐酸钾浓度增加幼苗株高、茎粗、干鲜重均有不同程度的增加,表明黄腐酸钾一定程度上缓解了干旱低温胁迫对西葫芦幼苗生长的抑制。 这与周海涛[18] 、王研[19] 等的研究结果基本一致。植物可以通过渗透调节物质脯氨酸、可溶性蛋白、可溶性糖的累积增加细胞液浓度和降低水势,以增强吸水能力,抵御逆境造成的危害[20] 。本研究中,黄腐酸钾处理一定程度上提高干旱低温胁迫下西葫芦幼苗叶片的脯氨酸、可溶性糖、可溶性蛋白含量。 这与赵永长[9] 、孙海燕[21] 等的研究结果部分一致,表明黄腐酸钾可以增加干旱低温胁迫下西葫芦幼苗的细胞液浓度,调节细胞渗透压,提高幼苗的抗逆性。
受到非生物胁迫时,植物体内活性氧会积累,加重膜脂过氧化致使膜系统受损,其主要产物丙二醛直接反映细胞膜过氧化程度,间接反映植物体内活性氧水平[22-24] 。 植物体内的SOD、POD、CAT 是重要的抗氧化酶,可以清除超氧阴离子自由基、过氧化氢和羟自由基,减少活性氧,降低膜脂过氧化,稳定细胞膜透性[25-27] 。 本研究表明,干旱低温胁迫下西葫芦幼苗叶片的MDA 含量增加,SOD、POD、CAT 活性升高,这与前人研究结果基本一致。 张小冰等[28] 研究发现,腐植酸提高干旱胁迫下玉米幼苗叶片的SOD、POD、CAT 活性,降低MDA 含量。 金平等[29] 研究表明,低温胁迫下腐植酸可提高水稻叶片脯氨酸含量,减少MDA含量,提高水稻抗冷性。 本试验结果表明,黄腐酸钾一定程度上提高干旱低温胁迫下西葫芦幼苗叶片的SOD、POD、CAT 活性,降低其MDA 含量。说明黄腐酸钾能够减轻膜脂过氧化程度,提高植物的抗逆性。
叶绿素是植物进行光合作用的主要色素,在一定范围内光合速率与叶绿素含量关系密切[30,31] 。 本试验中,干旱低温胁迫一定程度上减少幼苗叶片中类胡萝卜素和叶绿素的合成。 其原因可能是,该胁迫导致光合色素降解程度加重,从而使叶绿素积累量下降[32] ,而黄腐酸钾能够抑制色素降解。 植物在非生物胁迫下可以通过光合参数Pn、Ci、Gs、Tr 的变化反映光合作用的强弱[33] 。李英浩等[34] 研究表明,腐植酸可以改善干旱胁迫下燕麦叶片的光合性能;张彩凤等[12] 研究指出,腐植酸钾可以减轻低温胁迫后植物的蒸腾速率、气孔导度、净光合速率的降低程度。 本试验结果表明,干旱低温胁迫下西葫芦幼苗叶片的Pn、Ci、Gs、Tr 显著降低,而干旱低温胁迫下黄腐酸钾处理西葫芦幼苗叶片的各光合参数值与DL 相比显著增加。 这说明黄腐酸钾能够增强干旱低温胁迫下西葫芦幼苗叶片光合色素的稳定性,提高光合速率。
叶绿素荧光参数是研究植物光合功能的重要指标,可以反映植物在逆境下光合作用的真实行为[35] 。 本试验中,干旱低温胁迫下西葫芦幼苗叶片的Fv/ Fm 、ΦPSⅡ、ETR、qP 低于对照(CK),Fo、NPQ 则高于CK。 这表明西葫芦幼苗叶片PSⅡ光合机构受到伤害,热能形式消耗掉的能量增加,光合效率降低,与杜清洁[36] 、卞凤娥[37] 等的研究结果类似。 干旱低温胁迫下黄腐酸钾处理西葫芦幼苗叶片的Fv/ Fm、ΦPSⅡ、ETR、qP 均有不同程度的升高,Fo、NPQ 則降低,表明黄腐酸钾在一定程度上缓解干旱低温胁迫对西葫芦幼苗叶片PSⅡ的伤害,提高PSⅡ反应中心的光化学活性,继而提高光合效率。
4 结论
本研究表明:黄腐酸钾一定程度上能够减轻干旱低温胁迫对西葫芦幼苗生长的抑制;提高抗氧化酶活性,降低活性氧含量;降低MDA 含量,减轻膜脂过氧化程度;增加渗透调节物质含量,维持细胞内渗透平衡;增加叶绿素含量,增强光合作用,降低PSⅡ光合机构受到的伤害,提高光合效率,增强西葫芦幼苗的抗逆性。 整体上看,各处理中以0.05%黄腐酸钾处理(T3)效果最佳,西葫芦幼苗叶面积、全株鲜重、全株干重、脯氨酸含量和SOD、POD、CAT 活性与DL(干旱低温胁迫)相比,分别显著增加13.89%、16.20%、17.43%、19.43%和13.48%、7.13%、16.28%,MDA 含量降低14.37%;叶绿素总量、净光合速率显著增加19. 66% 和17.95%;叶片的Fv/ Fm、ΦPSⅡ、ETR、qP 显著升高4.25%、17.57%、14.85%、14.01%,Fo、NPQ 分别降低5.30%、15.79%。 综上,黄腐酸钾可以在一定程度上缓解干旱低温胁迫对西葫芦幼苗生长和生理生化的抑制作用,以0.05%黄腐酸钾处理(T3)效果较为显著。
参 考 文 献:
[1] 杜社妮,白岗栓,梁银丽. 土壤水分和光照对西葫芦生长和生理特性的影响[J]. 应用生态学报,2011,22(4):1101-1106.
[2] 刘术均,刘爱群,何明. 外源甜菜碱对低温胁迫下茄子幼苗抗性的影响[J]. 北方园艺,2018(22):57-61.
[3] 谷端银. 腐植酸对氮胁迫下黄瓜生长及生理代谢的影响[D]. 泰安:山东农业大学,2016.
[4] 王晓宁,魏永义,周伟,等. 水杨酸对盐胁迫下黄瓜种子萌发和幼苗根系生长的影响[J]. 分子植物育种,2022,20(2):524-528.
[5] 王建霞,李昕,范楷,等. 海藻酸钠寡糖和赤霉素复配剂对亚低温胁迫后设施番茄生长发育的影响[J]. 山东农业科学,2022,54(7):122-128.
[6] 朱世琦,孙晓刚. 外源脱落酸对铅胁迫下王族海棠光合特性的影响[J]. 山东农业科学,2022,54(6):93-98.
[7] 柴文臣,阎世江. 甜菜碱对干旱胁迫下茄子幼苗生长及生理指标的影响[J]. 中国瓜菜,2021,34(8):78-83.
[8] 吴雪霞,朱宗文,张爱冬,等. 外源褪黑素对低温胁迫下茄子幼苗生长及其光合作用和抗氧化系统的影响[J]. 西北植物学报,2017,37(12):2427-2434.
[9] 赵永长. 黄腐酸钾对干旱胁迫下烤烟生长的调控效应及机理研究[D]. 北京:中国农业科学院,2017.
[10] 刘玉丰,闫征南,万泽,等. 盐碱胁迫下黄腐酸对甜瓜幼苗生长抑制的缓解效应[J]. 北方园艺,2021(24):45-50.
[11] 刘彩娟,吕春雨,艾希珍,等. 黄腐酸对干旱胁迫下黄瓜光合特性及产量和品质的影响[J]. 应用生态学报,2022,33(5):1300-1310.
[12] 张彩凤,王慧,潘虹,等. 低温胁迫下叶面喷施腐植酸钾对红掌生理生化指标的影响[J]. 山西农业科学,2015,43(2):167-171,191.
[13] 李建明,王平,李江. 灌溉量对亚低温下温室番茄生理生化与品质的影响[J]. 农业工程学报,2010,26(2):129-134.
[14] 董绪兵,毕焕改,刘业霞,等. 黄瓜幼苗干旱-低温交叉适应与渗透调节的关系[J]. 中国农业科学,2011,44(2):335-340.
[15] 毛丽萍,郭尚. 回归相关法测定西葫芦叶面积研究[J]. 上海蔬菜,2008(5):74-75.
[16] 高俊凤,孙群,曹翠玲,等. 植物生理学实验指导[M]. 北京:高等教育出版社,2006.
[17] 王新军,阎世江. 干旱胁迫对番茄幼苗生理特性的影响[J].中国瓜菜,2022,35(6):76-80.
[18] 周海涛,张艳阳,赵孟圆,等. S-诱抗素和黄腐酸对干旱胁迫下皮燕麦抗旱生理特性的影响[J]. 农学学报,2020,10(4):7-13.
[19] 王研,李灵芝,郭文忠,等. 腐植酸对西葫芦幼苗生长、生理及水分利用率的影响[J]. 中国农学通报,2022,38(13):47-53.
[20] 李怀伟. 旱涝-低温交叉胁迫对小麦抗寒性的影响[D]. 哈尔滨:东北农业大学,2013.
[21] 孙海燕,孔德庸,胡慧影,等. 腐植酸浸种对低温胁迫下玉米幼苗抗氧化系统的影响[J]. 生态学报,2021,41(13):5385-5397.
[22] Kar R K. Plant responses to water stress:role of reactive oxygenspecies[J]. Plant Signaling & Behavior,2011,6(11):1741-1745.
[23] Lawlor D W,Cornic G. Photosynthetic carbon assimilation andassociated metabolism in relation to water deficits in higherplants[J]. Plant,Cell & Environment,2002,25(2):275-294.
[24] Yuan G F,Jia C G,Li Z,et al. Effect of brassinosteroids ondrought resistance and abscisic acid concentration in tomato un ̄der water stress[J]. Scientia Horticulturae,2010,126(2):103-108.
[25] 王寶山. 生物自由基与植物膜伤害[J]. 植物生理学通讯,1988(2):12-16.
[26] 蒋明义,郭绍川. 水分亏缺诱导的氧化胁迫和植物的抗氧化作用[J]. 植物生理学通讯,1996(2):144-150.
[27] 朱维琴,吴良欢,陶勤南. 作物根系对干旱胁迫逆境的适应性研究进展[J]. 土壤与环境,2002,11(4):430-433.
[28] 张小冰,邢勇,郭乐,等. 腐植酸钾浸种对干旱胁迫下玉米幼苗保护酶活性及MDA 含量的影响[J]. 中国农学通报,2011,27(7):69-72.
[29] 金平,刘山莉. 腐殖酸与水稻抗冷性的研究初探[J]. 东北农业大学学报,1997,28(1):90-93.
[30] Lin Y P,Lee T Y,Tanaka A,et al. Analysis of an Arabidopsisheat ̄sensitive mutant reveals that chlorophyll synthase is in ̄volved in reutilization of chlorophyllide during chlorophyll turn ̄over[J]. The Plant Journal,2014,80(1):14-26.
[31] 李岩,田维丽,谢恩俊,等. 低温胁迫及生理生化指标综合评价抗寒茶树[J]. 种子,2020,39(12):38-43,54.
[32] Santos C V. Regulation of chlorophyll biosynthesis and degrada ̄tion by salt stress in sunflower leaves[J]. Scientia Horticultu ̄rae,2004,103(1):93-99.
[33] 隋昌成. 外源亚精胺对葡萄幼苗盐碱胁迫的缓解效应[D].泰安:山东农业大学,2021.
[34] 李英浩,刘景辉,朱珊珊,等. 干旱胁迫下腐植酸对燕麦叶片光合性能的调控效应[J]. 麦类作物学报,2019,39(11):1385-1391.
[35] Salvatori E,Fusaro L,Gottardini E,et al. Plant stress analysis:application of prompt,delayed chlorophyll fluorescence and 820nm modulated reflectance. Insights from independent experi ̄ments[J]. Plant Physiology and Biochemistry,2014,85:105-113.
[36] 杜清洁,代侃韧,李建明,等. 亚低温与干旱胁迫对番茄叶片光合及荧光动力学特性的影响[ J]. 应用生态学报,2015,26(6):1687-1694.
[37] 卞凤娥,唐翠花,邢浩,等. 外源褪黑素对干旱胁迫下葡萄内源褪黑素及叶绿素荧光特性的影响[J]. 植物生理学报,2018,54(10):1615-1623.
