外源水杨酸对铬胁迫下燕麦种子萌发和幼苗生理特性的影响
2024-04-01王志科唐慧玲邓金霞杨澳红
王志科 唐慧玲 邓金霞 杨澳红
摘要:为了探究外源水杨酸(SA)对铬(Cr6+ )胁迫下燕麦(Avena sativa L.)种子萌发抑制的缓解效应,本试验采用水培法,首先筛选出100 mg/ L 为适宜的镉胁迫处理浓度,然后设置不同浓度(0.01、0.02、0.03、0.04、0.08、0.10 mmol/ L)外源SA 浸种处理,以蒸馏水浸种为对照(CK),研究SA 对100 mg/ L 铬胁迫下燕麦种子萌发、幼苗生长、抗氧化酶的活性和渗透调节物质及MDA 含量的影响。 结果表明,100 mg/ L Cr6+ 溶液胁迫下燕麦种子的萌发和幼苗生长均被显著抑制;外源SA 浸种处理能够通过激活燕麦幼苗CAT、POD 和SOD 等抗氧化酶活性,减少MDA 含量,提高可溶性糖和脯氨酸含量,来缓解重金属铬对燕麦种子萌发的毒害作用,进而增强燕麦幼苗的抗逆性。 本试验条件下,以0.03 mmol/ L 外源SA 浸种处理的效果较优,明显缓解Cr6+ 胁迫对燕麦种子萌发、幼苗生长及相关生理指标的抑制。
关键词:水杨酸;铬胁迫;燕麦;种子萌发;幼苗生长;生理特性
中图分类号:S512.6文献标识号:A文章编号:1001-4942(2024)01-0091-06
我国耕地土壤中重金属铬浓度较高[1] ,且土壤中Cr6+ 的点位超标率为1.1%[2] 。 过量的铬不仅损害植物及土壤, 还会对人类健康造成严重影响[3] 。 重金属的积累不仅取决于植物对重金属的吸收能力, 还与土壤中的生物密切相关[4] 。 土壤中铬主要以Cr6+ 和Cr3+ 两种形态存在,不同形态的铬对植物的毒害程度不同;土壤固定Cr3+ 的能力显著高于Cr6+,Cr6+ 的生物毒性显著高于Cr3+, 土壤理化性质对铬的生物毒性影响较显著[5] 。 因此,研究如何缓解重金属铬对植物的毒害作用是一項重要课题。
水杨酸(salicylic acid, SA)是一种脂溶性有机酸,白色结晶粉状物,是植物体内普遍存在的一种小分子酚类物质;SA 作为一种信号分子参与植物多种非生物胁迫过程,提高植物对逆境的抵抗能力[6] 。 研究表明,外源水杨酸能缓解三嗪类除草剂对玉米幼苗的毒害作用[7] 。 施加100 μmol/ L外源水杨酸能显著缓解镉胁迫对藜麦幼苗生长的抑制作用[8] 。 1 mmol/ L 外源水杨酸可显著提高低盐胁迫下草木樨种子的萌发能力和促进幼苗生长[9] 。 0.25 mmol/ L 外源水杨酸浸种能促进月见草种子萌发[10] 。
燕麦(Avena sativa L.)不耐高温,多于阴凉环境中生长,但光照不足会造成其发育不良,影响产量。 近年来,重金属污染土壤导致燕麦的萌发和生长受到明显抑制,其产量下降[11] 。 目前关于外源SA 缓解Cr6+胁迫对燕麦影响方面的研究鲜有报道。 本试验研究不同浓度外源SA 对铬胁迫下燕麦种子萌发及幼苗生理特性的影响,以期为提高其耐铬胁迫能力和产量提供参考。
1 材料与方法
1.1 试验材料
燕麦种子由陇东学院生命科学与技术学院植物生理实验室提供。 三氧化二铬(Cr2 O3 )、水杨酸、磷酸二氢钾、磷酸氢二钾、30% H2 O2、愈创木酚、L-甲硫氨酸、氯化硝基四氮唑蓝(NBT)、核黄素均购自生工生物工程(上海)股份有限公司。
1.2 试验设计
试验于2022 年4—8 月在陇东学院生命科学与技术学院植物生理实验室进行。
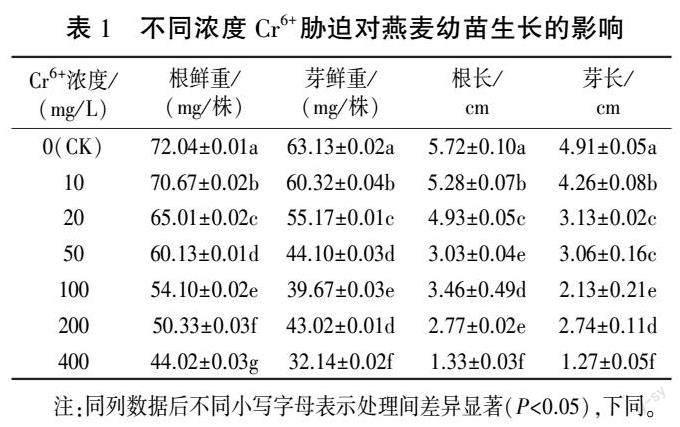
1.2.1 Cr6+胁迫浓度筛选试验 挑选大小均一且饱满的燕麦种子,用5% NaClO 溶液消毒10 min后,用蒸馏水浸洗2~3 次,洗去种子表面残留的NaClO 溶液。 将种子放入铺有双层滤纸的培养皿中,每皿50 粒,分别用10、20、50、100、200、400mg/ L 的Cr6+溶液10 mL 处理燕麦种子,以蒸馏水为对照(CK),共7 个处理。 每个处理重复3 次。将所有培养皿放入(25±1) ℃、相对湿度85% ~90%的恒温培养箱中培养7 d。 在此期间,每天固定时间统计发芽种子数,定时补充相应浓度的Cr6+溶液。 种子萌发稳定后,测量芽长、根长、芽鲜重和根鲜重,计算发芽相关指标。
1.2.2 外源SA 浸种缓解Cr6+ 胁迫试验 根据
1.2.1试验结果得出适宜铬胁迫浓度为100 mg/ L后,用该浓度Cr6+ 溶液浸种处理试验材料。 挑选大小均一且饱满的燕麦种子,用5% NaClO 溶液消毒10 min 后,用蒸馏水浸洗2~3 次,洗去种子表面残留的NaClO 溶液。 将种子放入小烧杯中,每杯50 粒,分别用0.01、0.02、0.03、0.04、0.08、0.10 mmol/ L SA 溶液20 mL 浸种8 h 后,用1.2.1中的方法进行100 mg/ L Cr6+ 溶液胁迫处理,同时用等量蒸馏水和100 mg/ L Cr6+ 溶液浸种作为空白对照和条件对照,共8 个处理。 具体如下:蒸馏水浸种( CK)、100 mg/ L Cr6+ 浸种胁迫处理( Cr6+ )、0. 01 mmol/ L SA 浸种Cr6+ 胁迫处理(0.01SA+Cr6+)、0.02 mmol/ L SA 浸种Cr6+ 胁迫处理(0.02SA+Cr6+)、0.03 mmol/ L SA 浸种Cr6+ 胁迫处理(0.03SA+Cr6+)、0.04 mmol/ L SA 浸种Cr6+ 胁迫处理(0.04SA+Cr6+)、0.08 mmol/ L SA 浸种Cr6+胁迫处理(0.08SA+Cr6+ )和0.10 mmol/ L SA 浸种Cr6+胁迫处理(0.10SA+Cr6+ )。 每个处理重复3次。 浸种结束后,用滤纸吸干种子上的残留溶液,将种子放入铺有双层滤纸的培养皿中,每皿50粒,共8 个处理组,最后将所有培养皿放入(25±1) ℃、相对湿度85%~90% 的恒温培养箱中培养7 d。 在此期间,每天固定时间统计发芽种子数,定时补充相应浓度的Cr6+ 溶液。 种子萌发稳定后,测量芽长、根长、芽鲜重和根鲜重,计算发芽相关指标。
1.3 测定项目及方法
1.3.1 形态指标 种子发芽以胚芽长为种子长度一半为发芽标准。 在50 粒试验种子中随机选取15 粒测量芽长、根长并取平均值。 燕麦种子的发芽率、发芽势、发芽指数、活力指数的计算参照李剑[12] 的方法进行。
1.3.2 生理指标 CAT、SOD 活性参照李剑[12] 的方法测定;POD 活性参照程艳等[13] 的方法测定;MDA 含量参照赵炀等[14] 的方法测定;脯氨酸和可溶性糖含量参照李丽等[15] 的方法测定。
1.4 数据处理与分析
用Microsoft Excel 2016 进行数据处理, 用SPSS 26.0 进行单因素方差分析。 表中数据为平均值±标准差。
2 结果与分析
2.1 Cr6+胁迫对燕麦幼苗生长的影响
从表1 中可以看出,随着Cr6+ 胁迫浓度的升高,燕麦幼苗根鲜重、芽鲜重、根长和芽长均被显著抑制。 与CK 相比, 在10、20、50、100、200、400mg/ L Cr6+胁迫下,燕麦幼苗的根鲜重分别显著降低1. 90%、9. 76%、16. 53%、24. 90%、30. 14%、38.90%,芽鲜重分别显著降低4.45%、12.61%、30.14%、37.16%、31.85%、49.09%,根长分别显著减小7.69%、13.81%、47.03%、39.51%、51.57%、76.75%,芽长分别显著减小13. 24%、36. 25%、37.68%、56.62%、44.20%、74.13%。
2.2 Cr6+胁迫对燕麦种子萌发的影响及胁迫浓度筛选
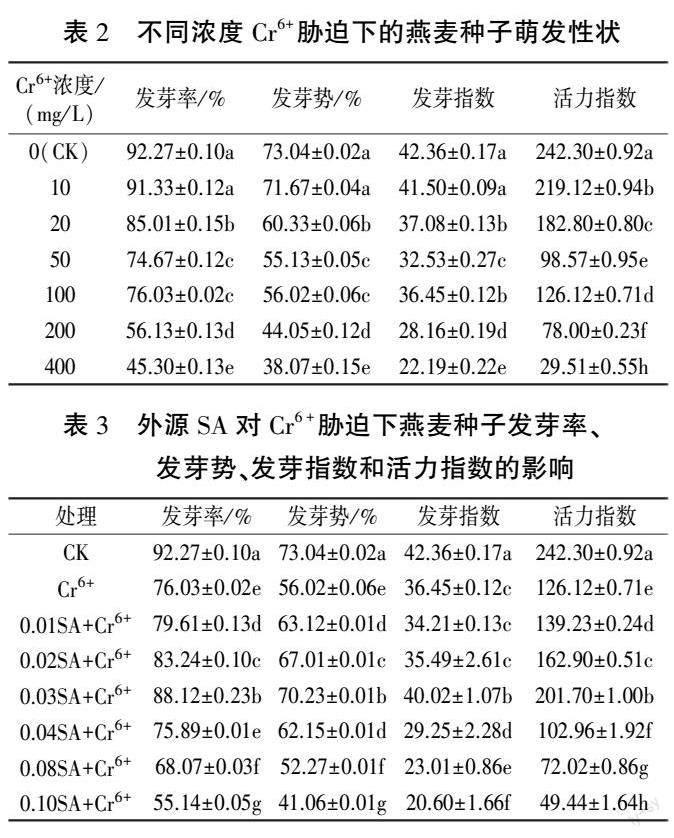
从表2 中可以看出,在不同浓度Cr6+ 胁迫下,燕麦种子萌发受到不同程度的抑制。 在10 mg/ LCr6+胁迫下,燕麦种子的发芽率、发芽势和发芽指数都下降,但与对照组差异不显著;在20、50、100、200 mg/ L 和400 mg/ L Cr6+胁迫下,燕麦种子的这三个指标均显著下降。 不同浓度Cr6+ 胁迫下,燕麦种子的活力指数均显著下降。 结合表1结果来看,100 mg/ L Cr6+胁迫处理下,燕麦种子的发芽率、发芽势、发芽指数、活力指数均显著下降,幼苗生长被显著抑制,且对活力指数的抑制程度约为50%。 因此,本试验选用100 mg/ L Cr6+ 溶液作为燕麦种子Cr6+胁迫的处理浓度。
2.3 外源SA 对Cr6+胁迫下燕麦种子萌发的影响
从表3 中可以看出,与CK 相比,Cr6+ 胁迫下燕麦种子发芽率、发芽势、发芽指数、活力指数分别显著降低17.60%、23.30%、13.95%、47.95%;与Cr6+处理相比,0.03SA+Cr6+ 处理显著缓减Cr6+ 胁迫对燕麦种子萌发的抑制,发芽率、发芽势、发芽指数、活力指数分别显著提高15.90%、25.37%、9.79%、59.93%。 表明适宜浓度的外源SA 能显著缓减Cr6+胁迫对燕麦种子萌发的抑制。
2.4 外源SA 对Cr6+胁迫下燕麦幼苗生长的影响
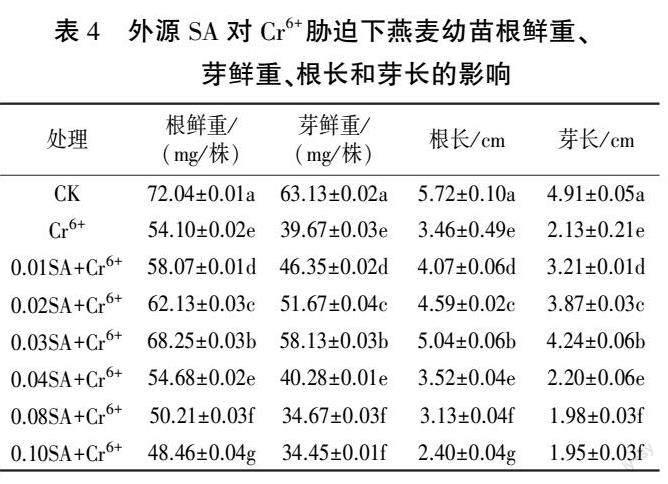
从表4 中可以看出,相比于CK,Cr6+ 胁迫下燕麦幼苗的根鲜重、芽鲜重、根长、芽长分别显著减少24.90%、37.16%、39.51%、56.62%。 与Cr6+处理相比,Cr6+胁迫下外源施加不同浓度SA 处理中,0.03SA+Cr6+处理燕麦幼苗的根鲜重、芽鲜重、根长、芽长值均最大,仅次于CK(P <0.05),较Cr6+处理分别增26.16%、46.53%、45.66%、99.06%(P<0.05)。
2.5 外源SA 对Cr6+胁迫下燕麦幼苗抗氧化酶活性的影响
从表5 中可以看出,与CK 相比,Cr6+ 胁迫处理显著降低燕麦幼苗根、芽中CAT、POD、SOD 活性。 Cr6+胁迫下外源施加不同浓度SA 处理中,0.03SA+Cr6+处理燕麦幼苗的根、芽中这三种酶活性最高,仅次于CK;与Cr6+ 处理相比,0.03SA+Cr6+ 处理幼苗根、芽中CAT 活性分別显著提高57.04%、26.03%, POD 活性分别显著提高15.17%、16.49%,SOD 活性分别显著提高32.20%、28.25%。 0.1 mmol/ L SA 处理不能缓减燕麦幼苗根和芽中三种酶活性的下降,与Cr6+ 处理相比反而加大了这三种酶活性的下降幅度。 表明适宜浓度的外源SA 能缓减Cr6+ 胁迫对燕麦幼苗造成的伤害,高浓度SA 反而会加重Cr6+胁迫对燕麦幼苗的伤害。
2.6 外源SA 对Cr6+ 胁迫下燕麦幼苗MDA、脯氨酸、可溶性糖含量的影响
MDA 是脂膜过氧化的主要产物。 表6 结果表明,Cr6+ 胁迫导致燕麦幼苗根、芽中MDA 含量较CK 显著增加,外源SA 浸种处理则显著降低Cr6+胁迫下根、芽中MDA 含量的增加幅度,且随着外源SA 浓度增加降低效果呈先增加后降低的变化。 与Cr6+ 处理相比,不同浓度外源SA 处理中,0.03SA+Cr6+处理幼苗根、芽中的MDA 含量均最低,分别显著降低32.30%、26.11%。
脯氨酸是植物蛋白质的组成成分之一,在植物中以游离态形式广泛存在。 植物组织中脯氨酸含量增加,可提高植物的抗逆性。 表6 结果表明,与CK 相比,Cr6+ 处理燕麦幼苗根、芽中脯氨酸含量显著增加;外源SA 浸种处理则显著提高Cr6+胁迫下根、芽中脯氨酸含量,且随着外源SA 浓度增加其增幅呈先升后降的变化;与Cr6+ 处理相比,0.03SA+Cr6+ 处理幼苗的根、芽中脯氨酸含量最高,分别显著提高81.98%、91.27%。
可溶性糖在降低植物细胞渗透势、维持细胞膜结构等方面都具有重要作用。 表6 结果表明,Cr6+处理下燕麦幼苗根、芽中可溶性糖含量较CK显著升高。 与Cr6+处理相比,外源SA 浸种处理显著提高Cr6+胁迫下根、芽中可溶性糖含量,且随着外源SA 浓度增加其增幅呈先升后降的变化;与Cr6+处理相比,0.03SA+Cr6+处理幼苗根、芽中可溶性糖含量最高,分别显著增加110.31%、56.68%。
表明外源SA 浸种处理能显著增加Cr6+ 胁迫条件下燕麦幼苗根、芽中可溶性糖含量,有效降低燕麦幼苗组织细胞渗透势,从而减缓Cr6+ 胁迫对燕麦幼苗的伤害。
3 讨论
土壤重金属污染会导致农田用地减少、粮食产量降低,可给当地经济造成极大的损失[16] 。 有研究表明:铬胁迫明显抑制黄瓜种子的萌发[17] ;低浓度Cr3+ 促进茼蒿种子的萌发,高浓度Cr3+ 抑制其萌发[18] ;低浓度Cr6+ 促进苜蓿种子的萌发,高浓度Cr6+抑制其萌发[19] ;Cr6+ 胁迫显著抑制百日草[20] 和娃娃菜[21] 种子的萌发和幼苗生长。 本研究表明,100 mg/ L Cr6+胁迫处理下,燕麦种子的发芽率、发芽势、发芽指数和活力指数均显著降低,这与前人研究结果一致;同时,燕麦幼苗的根鲜重、芽鲜重、根长和芽长也均显著降低。
水杨酸(SA)作为一种小分子酚类化合物,具有多种生理调控作用[22] 。 研究表明:SA 可以促进NaCl 胁迫下大豆种子萌发[23] ,提高铬胁迫下绿豆幼苗的生长[24] ,还可以缓解Cr(Ⅵ)对玉米生长的抑制作用[25] ,并能缓解低温胁迫对菜豆种子萌发的抑制[26] 。 本研究表明,0.03 mmol/ L SA处理显著缓减Cr6+胁迫对燕麦种子萌发及幼苗根鲜重、芽鲜重、根长和芽长的抑制,这与前人研究结果一致。
重金属胁迫会引起植物细胞产生大量活性氧(ROS),对细胞膜造成氧化损伤[27] 。 有研究表明,铬胁迫能增加小麦幼苗中丙二醛含量,使其生长受到严重损伤[28] 。 本研究表明,Cr6+ 胁迫导致燕麦幼苗根、芽中MDA 含量显著增加,适宜浓度外源SA 浸种处理显著降低Cr6+ 胁迫下其含量。表明外源SA 浸种能缓减铬胁迫造成的燕麦幼苗氧化损伤。 正常情况下,植物体内ROS 的产生与清除处于平衡状态,当其受到重金属胁迫时,会打破此平衡,植物会激活CAT、POD、SOD 等抗氧化酶活性来清除过多的ROS。 潘法康等[29] 研究表明,Cr6+胁迫使薄荷幼苗中CAT、SOD 活性呈现先下降后上升趋势,POD 活性则呈先升高后降低趋势。 本研究表明,Cr6+ 胁迫处理显著降低燕麦幼苗根、芽CAT、POD、SOD 活性,0.03 mmol/ L SA 浸种处理显著缓减三种酶活性的下降幅度。 表明适宜浓度外源SA 浸种可能通过提高幼苗抗氧化酶活性来缓减Cr6+胁迫对燕麦造成的伤害。
脯氨酸和可溶性糖是参与植物渗透调节的重要物质[30] 。 已有研究表明,铬胁迫能诱导紫花苜蓿[19] 、玉米[30] 、白三叶[31] 、茳芏[32] 中脯氨酸含量显著增加。 本研究表明,0.03 mmol/ L SA 浸种处理显著提高Cr6+ 胁迫下幼苗根、芽中脯氨酸和可溶性糖含量。 表明适宜浓度外源SA 浸种可能通过提高幼苗根、芽中脯氨酸和可溶性糖含量来缓减Cr6+胁迫对燕麦造成的伤害。
4 结论
本研究得出,外源SA 浸種可能通过激活燕麦幼苗CAT、POD、SOD 活性,减少MDA 含量和提高可溶性糖、脯氨酸含量等来缓解铬胁迫对燕麦种子萌发和幼苗生长的毒害作用,进而增强燕麦幼苗的抗逆性,且以0.03 mmol/ L 外源SA 浸种处理效果最优。
参 考 文 献:
[1] 曹俊雅,张婧,张文茜,等. 土壤中重金属铬(Ⅵ)污染修复技术的研究进展[J]. 土壤通报,2022,53(5): 1220-1227.
[2] 徐国豪,刘英豪,常明慧,等. 土壤外源铬的作物毒性响应及富集差异研究[J]. 农业环境科学学报,2023,42(2):284-290.
[3] 白雪,赵鑫宇,景秀清,等. 农田土壤中真菌群落在时间序列对铬胁迫的响应机制[J]. 中国生态农业学报:中英文,2022,30(1): 105-115.
[4] 蔡苗苗. 硒对铬污染土壤上小白菜生长与铬吸收的调节及其根际过程研究[D]. 武汉:华中农业大学,2021.
[5] 付平南,贡晓飞,罗丽韵,等. 不同价态铬和土壤理化性质对大麦根系毒性阈值的影响[J]. 环境科学,2020,41(5):2398-2405.
[6] 宋雅娟,李师翁. 水杨酸在植物缓解重金属镉胁迫中的作用[J]. 环境科学与技术,2020,43(2): 177-186.
[7] 余倩倩. 外源水杨酸缓解莠去津、特丁津和莠灭净对玉米毒性的作用[D]. 南京:南京农业大学,2019.
[8] 杨张钰,朱心雨,胡乐佳,等. 外源水杨酸对镉胁迫下藜麦幼苗生长生理的缓解效应[J]. 江苏农业科学,2022,50(4): 52-57.
[9] 刘泓言,任德,向春阳. 水杨酸对盐胁迫下草木樨种子萌发和幼苗生长的影响[J]. 天津农林科技,2022(1): 18-20.
[10] 姜云天,张家兴,王铭宇,等. 外源水杨酸对长白山区月见草种子萌发的影响[J]. 园艺与种苗,2022,42(11): 1-3,21.
[11] 闫超凡,叶丽丽,陈余道,等. 多金属胁迫对燕麦重金属累积及生理特征的影响[J]. 桂林理工大学学报,2020,40(3): 597-605.
[12] 李剑. 成熟度和粒位对大麦种子活力和耐藏力影响的研究[D]. 石河子:石河子大学,2020.
[13] 程艳,陈璐,米艳华,等. 水稻抗氧化酶活性测定方法的比较研究[J]. 江西农业学报,2018,30(2): 108-111.
[14] 赵炀,李永生,高秀峰. 基于愈创木酚荧光减量准确测定过氧化物酶活性的新方法[J]. 分析化学,2015,43(7): 1040-1046.
[15] 李丽,尹欣茹. 不同浓度氯化镉对碱蓬幼苗组织生理生化指标的影响[J]. 中国林副特产,2022(4): 16-18.
[16] 穆海婷,王英哲,苗一凡,等. 重金属铜和铅胁迫对东方山羊豆幼苗生长及生理特性的影响[J]. 草业学报,2022,31(11): 139-146.
[17] 张盼盼,纪江涛,武熊,等. 不同浓度EDTA 对铬胁迫下黄瓜种子萌发的缓解效应[J]. 榆林学院学报,2021,31(6): 47-52.
[18] 王小平. 铬胁迫对茼蒿种子萌发及幼苗抗逆生理影响的探究[J]. 南京晓庄学院学报,2017,33(6): 100-103.
[19] 刘长风,王椰,段士鑫,等. 铬胁迫下紫花苜蓿种子萌发和幼苗生理响应的主成分分析[J]. 江苏农业科学,2022,50(19): 176-182.
[20] 朱红霞,李赛楠,苏文青,等. 外源褪黑素对铬胁迫下百日草种子萌发及幼苗生理特性的影响[J]. 山东农业科学,2022,54(9): 64-69.
[21] 闫志强,陈银萍,余沛东,等. 外源一氧化氮对铬胁迫下娃娃菜种子萌发及幼苗生理特性的影响[J]. 西南农业学报,2019,32(9): 2033-2039.
[22] 白生文,张喜峰,李彩霞,等. 水杨酸在植物非生物胁迫中的作用研究进展[J]. 河西学院学报,2015,31(2): 19-24.
[23] 廖姝,倪祥银,齐泽民,等. 水杨酸对NaCl 胁迫下大豆种子萌发和幼苗逆境生理的影响[J]. 内江师范学院学报,2013,28(2): 39-42.
[24] 张小梅,王智鑫,李广录. 水杨酸对铬胁迫下绿豆幼苗生長抑制的缓解效应[J]. 河南农业科学,2012,41(11): 52-54.
[25] 贾亚宁. 水杨酸对铬胁迫下玉米幼苗脯氨酸含量影响实验研究[J]. 生物灾害科学,2017,40(2): 100-103.
[26] 孙梦笛,刘畅,冯国军,等. 水杨酸对低温胁迫下菜豆种子萌发及抗寒性的影响[J]. 黑龙江大学工程学报,2022,13(2): 91-96.
[27] 覃勇荣,赵贞涛,刘旭辉,等. 重金属胁迫对不同种源任豆种子萌发及幼苗生长的影响[J]. 江苏农业科学,2022,50(12): 156-162.
[28] 王燕,葛健文,牟伊,等. 铬胁迫下异甜菊醇浸种对小麦幼苗生长的影响[J]. 山东农业大学学报(自然科学版),2022,53(5): 711-718.
[29] 潘法康,张瑾,王楚石,等. 重金属铬胁迫对薄荷生长及生理特性的影响[J]. 安徽建筑大学学报,2020,28(3): 38-41.
[30] 葛春辉,张云舒,唐光木. 生物炭的施入对Cr(Ⅵ)胁迫条件下玉米幼苗的生理响应及铬吸收的影响[J]. 中国农学通报,2020,36(27): 20-25.
[31] 王铎,田雨龙,张鸿建,等. 胺鲜酯对铬胁迫下白三叶种子萌发特性的影响[J]. 草业科学,2021,38(10): 1986-1997.
[32] 韦江玲,潘良浩,陈元松,等. 重金属Cr6+ 胁迫对茳芏生理生态特征的影响[J]. 广西植物,2014,34(1): 89-94.
