胆固醇7α-羟化酶在毕赤酵母中的异源表达
2023-11-23赵昕杜玉瑶殷子扬毛淑红
赵昕 杜玉瑶 殷子扬 毛淑红
(1. 天津科技大学生物工程学院,天津 300457;2. 工业发酵微生物教育部重点实验室,天津 300457)
胆固醇7α-羟化酶(CYP7A1)又称细胞色素P450 7A1酶,是肝脏特异性微粒体细胞色素P450家族7亚家族A成员[1]。在肝脏中,CYP7A1催化胆固醇转化为7α-羟基胆固醇,从而促进胆汁酸的分解,CYP7A1为该反应的限速酶[2],对维持体内胆固醇代谢平衡起到关键的作用,其在保障胆固醇正常摄取、运输和转化方面有极其重要的作用。在体内,若胆固醇代谢异常易造成血管内壁的胆固醇沉积,形成动脉粥样硬化、胆结石等疾病,还可导致胆囊癌、结直肠癌等疾病的发生[3-4]。
22例携带GJB3基因c.538 C>T受检者电话随访中,针对听力情况特别是尖叫等高频区域是否受损进行问询,所有携带c.538的受检者均表示听力正常但未行耳鼻喉科专业检查,其中包含1例c.538 C>T纯合。GJB3基因c.538 C>T早期研究认为是常染色体显性遗传模式的高频率听力损失[7],但后期对于GJB3基因变异是否致病尚存在争议[8],结合本次报道临床对于GJB3基因c.538C>T的咨询建议应谨慎。
CYP7A1依赖于氧化酶和还原酶之间特定的电子传递链进行7位羟基化反应[5]。CYP7A1电子传递机制如图1,NADPH提供的两个电子转移到CPR的黄素腺嘌呤二核苷酸(FAD)区域,FAD接受电子会引起CPR的构象变化,使FAD接近黄素单核苷酸(FMN)区域,之后FAD处的电子转移到FMN;随后CPR分子恢复到原来的构象,允许FMN与CYP7A1分子电荷结合位点相互作用[6-9],然后电子流向CYP7A1的血红素铁区域,通过Fe=O机制介导胆固醇羟基化反应[10],CYP7A1催化反应可表示为:NADPH+H++O2+胆固醇→NADP++H2O+7α-羟基胆固醇。
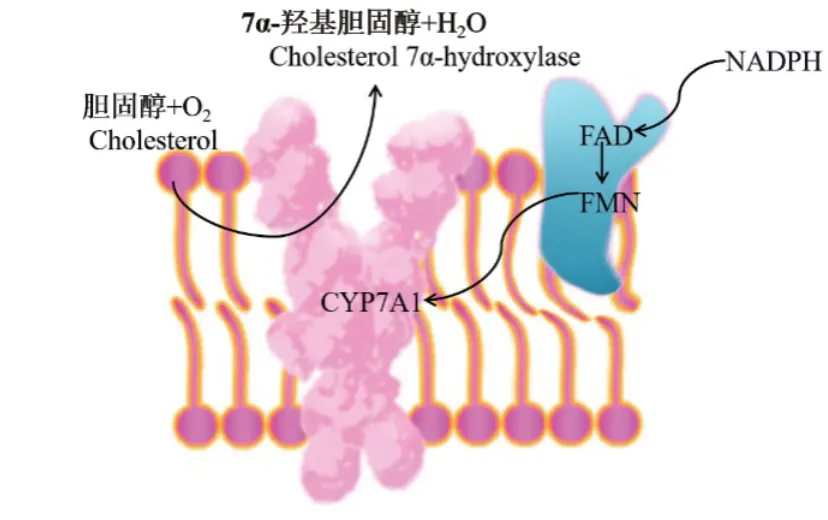
图1 胆固醇7α-羟化酶的电子传递机理Fig. 1 Electron transport mechanism of cholesterol 7α-hydroxylase
7α-羟基胆固醇不仅是一种胆固醇代谢产物,而且对胆汁酸合成、炎症和肿瘤等方面的治疗都有重要的作用。7α-羟基胆固醇在肝脏中被转化为胆汁酸,从而促进胆汁酸的合成和分泌[11]。此外,7α-羟基胆固醇可以减少炎性细胞浸润,降低炎症水平,从而起到抗炎作用[12]。最后,7α-羟基胆固醇可以抑制癌细胞的增殖和转移,具有一定的抗肿瘤作用[13]。因此构建一株利用CYP7A1高效生产7α-羟基胆固醇的菌株具有极大的医学价值和工业生产价值。
在构建表达胆固醇7α-羟化酶的毕赤酵母基因工程菌株的基础上,对7α-羟化酶进行理性设计与分子改造,对筛选获得的高活性7α-羟化酶适配不同来源CPR,并在毕赤酵母中共表达,从而为7α-羟基胆固醇高效生产奠定基础。
气相色谱-质谱法(GC‑MS)分析条件:升温程序:初始温度50℃,保持2 min,以5℃/min升到270℃,保持0 min,再以2.5℃/min升到290℃,保持0 min;再以10℃/min升到310℃,保持4 min,不分流进样,流速1 mL/min。
1 材料与方法
1.1 材料
限制性核酸内切酶(BamH I、EcoR I、Sal I和Sac I)、Goldstar best DNA聚合酶、Solution I连接酶、DNA提取纯化试剂盒和DNA凝胶回收试剂盒均为TaKaRa公司产品,胆固醇为Diamond公司产品,7α-羟基胆固醇为Genewiz公司产品。
1.2.5 胆固醇的转化 在YPD培养基上选取构建成功的重组菌株的单菌落,接种到30 mL YPG培养基中,30℃、220 r/min的摇床培养24 h;按培养基2%接种量接种到50 mL BMGY培养基中进行富集培养,30℃、220 r/min的摇床培养18 h,转接到BMMY培养基,于30℃、220 r/min的摇床继续培养24 h;加入0.01%底物胆固醇及1%诱导剂甲醇,继续培养4 d,期间每24 h添加甲醇至1%的浓度。取发酵液10 mL,并加入等体积乙酸乙酯轻微振荡萃取,离心后取上层有机相,浓缩至500 μL,使用薄层色谱(TLC)、气相色谱-质谱法(GC‑MS)及蒸发光分析产物的生成情况。
蒸发光检测分析条件:流动相为纯甲醇,进样量10 μL,载气流速2.2 mL/min,载气温度90℃。
只有肯在泥泞的小路上摸爬滚打,才有机会踏上铺满鲜花的大道。今年33岁的王振东,现任哈佳项目部总工程师。对于一个80后青年来说,这已经是值得称傲的成就了,但是踏遍工程现场的艰辛脚印向我们展示了他的成功当之无愧。
1.2 方法
1.2.1 重组质粒pPIC3.5K‑CYP7A1(WT/突变型)的构建 以合成的CYP7A1(WT)基因为模板,通过特异性引物(表1)PCR扩增克隆出WT基因及突变基因H111A、I114A、V281A、W284A和G485A,用限制性核酸内切酶BamH I和EcoR I对以上片段和pPIC3.5K质粒进行双酶切,切胶回收产物经Solution I连接酶过夜连接,将连接的产物化转至大肠杆菌E. coli JM 109,以卡那霉素抗性筛选重组转化子并测序。得到E. coli JM 109/pPIC3.5K‑CYP7A1(WT/突变型)。
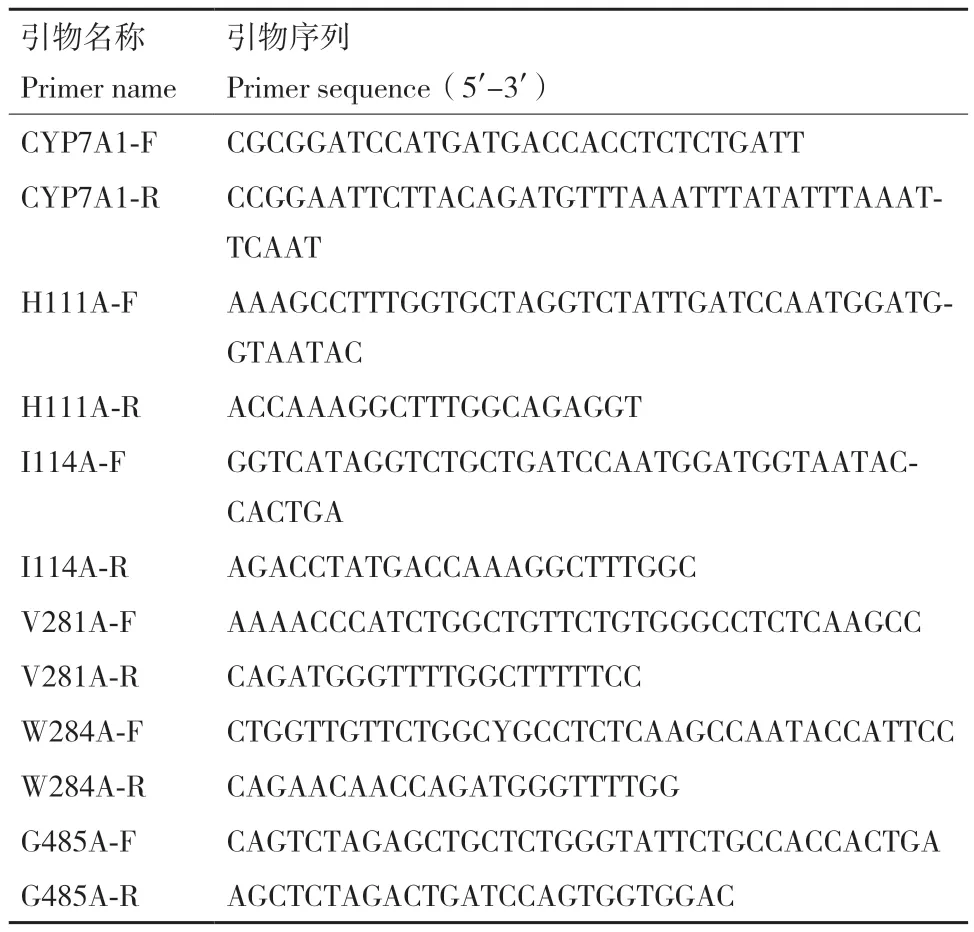
表1 CYP7A1基因野生型及突变型引物Table 1 Wild-type and mutant primers of CYP7A1 gene
1.2.2 毕赤酵母/pPIC3.5K‑CYP7A1(WT/突变型)菌株的构建 将测序正确的重组质粒pPIC3.5K‑CYP7A1(WT/突变型)用Sal I限制性核酸内切酶单酶切线性化;电转至毕赤酵母GS115感受态细胞中,使用含0.5 mg/mL G418的YPD和2.0 mg/mL G418的YPD培养基筛选高拷贝菌株,提取菌株基因组进行PCR验证,以获得重组菌株。
1.2.3 不同来源CPR与CYP7A1共表达毕赤酵母菌株的构建 将实验室保存的5种含不同来源CPR的质粒pPICZX‑CPR用Sac I限制性核酸内切酶单酶切线性化后电转至含CYP7A1(WT)基因的毕赤酵母感受态细胞中;5种CPR分别为人源氧化还原蛋白(hCPR)、酿酒酵母(Saccharomyces cerevisiae)来源氧化还原蛋白(ScCPR)、黑曲霉(Aspergillus nige)来源氧化还原蛋白(AnCPR)、大鼠来源氧化还原蛋白(RatCPR)及罗汉果来源氧化还原蛋白(SgCPR);使用含300 μg/mL博来霉素的YPDS和2.0 mg/mL G418的YPD培养基筛选高拷贝菌株,提取菌株基因组进行PCR验证,以获得重组菌株。
1.2.4 G485A与SgCPR共表达毕赤酵菌株的构建 将重组质粒pPICZX‑SgCPR用Sac I限制性核酸内切酶单酶切线性化,将线性化的质粒加入含CYP7A1(G485A)基因的毕赤酵母感受态细胞中,使用含300 μg/mL博来霉素YPDS和2.0 mg/mL G418的YPD培养基筛选高拷贝菌株,提取菌株基因组进行PCR验证,以获得重组菌株。
实际上,纵观近几年的邮品拍卖,只要是精品、珍品,就能引起买家的追逐,不仅成交相当活跃,而且市场潜力十分可观。如被称为我国“十大珍邮”之一的“全国山河一片红”邮票,单枚价格的涨势就十分惊人:早在1977年,就有150元一枚的身价;1987年,它的身价又涨至800元;到了1993年,成交价为3万元;在1997年,北京的一场艺术品拍卖会上,一枚的成交价高达17万元。2000年广州国际邮票钱币博览会上,一枚《全国山河一片红》邮票更是卖到18.5万元。据业内人士介绍,该邮票在拍卖场上,只要一亮相就深受藏家追捧,几乎没有流拍的事情发生。由此可见,只要投资者收藏有重大纪念事件方面的邮票,增值的可能性很大。
薄层色谱分析条件:展开剂为石油醚∶乙酸乙酯=2∶1,用含10%浓硫酸的无水乙醇喷洒TLC板,高温加热显色后,可清晰分辨胆固醇及产物7α-羟基胆固醇所在的位置。
师:同学们,老师觉得作者很了不起,广玉兰树形比较高大,我们平时看它只看到叶子,四季常青,感到冬季给我们带来生机。但是陈荒煤却不这样认为,他觉得藏在叶子里面的花更有生机。因此他着意来描写它。一株树上的花数世同堂,生生不息。有四种形态,其实作者也是有所侧重,写出了别人看不到,想不到的地方。你觉得他想突出哪种形态是最有生命力?
通过NCBI数据库获得全长人源CYP7A1基因(GenBank: NP_000771.2),由金唯智生物技术有限公司按照毕赤酵母密码子偏爱性进行优化合成。质粒pPIC3.5K(含有卡那霉素抗性基因)、pPICZX(含有博来霉素抗性基因)、大肠杆菌Escherichia coli JM 109、毕赤酵母GS115均由本实验室保存。
2 结果
2.1 重组毕赤酵母/pPIC3.5K‑CYP7A1对7α-羟基胆固醇产率的影响
通常,根据测量方法来划分,可以将地面三维激光扫描技术分成两种,分别是移动式及固定式激光扫描系统。[2]其中,固定式激光扫描系统,与传统测量中的全站仪是十分相似的,由内置数码相机、控制系统及激光扫描仪等多个部分组成。但是相对于全站仪来说,固定式激光扫描仪收集的是整个系列的“点云”数据信息,而不是分散的单点三维坐标,其主要特征是扫描面积大、速度较快、精准度较高以及具备良好的野外操作性。而移动式激光扫描系统,是以车载平台为基础,由惯性导航系统、全球定位系统与固定式三维激光扫描系统相结合而组成的。
薄层色谱定性分析结果(图2)显示,图中蓝色的样品点为7α-羟基胆固醇,粉色的样品点为胆固醇。毕赤酵母GS115即使在投底物胆固醇的情况下,也不会将胆固醇转化为7α-羟基胆固醇,而重组菌株具有催化胆固醇生成目的产物7α-羟基胆固醇的能力。
ADP受体拮抗剂噻吩并吡啶抑制ADP诱导的血小板聚集和活化,通过与P2Y型特殊巯基受体结合,抑制ADP受体的激活,常用于临床的有氯吡格雷、替格瑞洛和普拉格雷,有关专家在相关章节已经阐述。需要指出的是正在服用氯吡格雷患者,如准备进行CABG,除非血运重建紧急程度大于出血风险,拟行择期冠状动脉旁路移植术的患者,建议择期手术前停用氯吡格雷5天~7天。
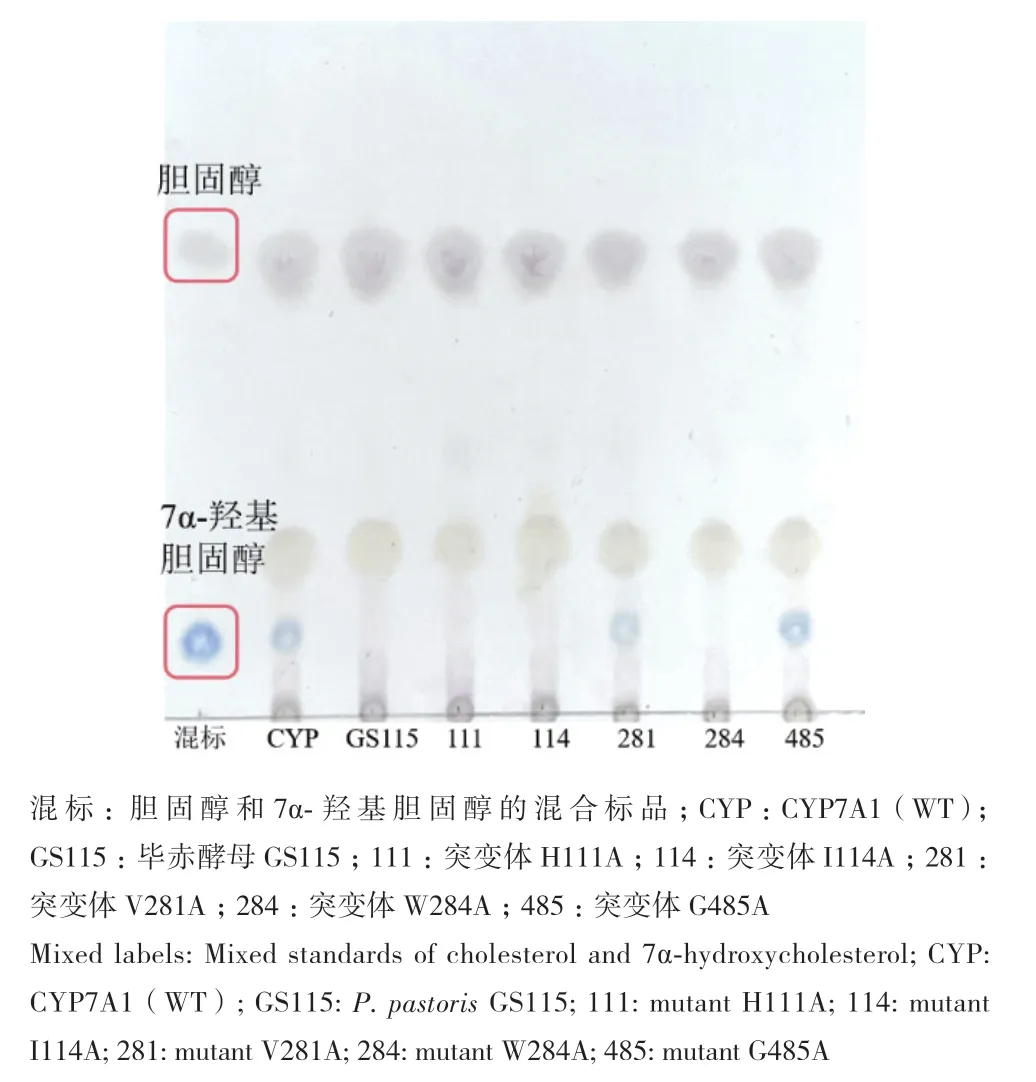
图2 重组毕赤酵母催化胆固醇TLC结果图Fig. 2 TLC analysis of biotransformation result of cholesterol catalyzed by recombinant P. pastoris
气相色谱-质谱法(GC‑MS)定性分析结果显示,图3‑A为7α-羟基胆固醇标品的质谱图,特征峰出现在m/z 456(分子离子峰)、129、95、73和57的位置。图3‑B为重组菌株转化产物的质谱图,特征峰与7α-羟基胆固醇标品的质谱图的特征峰相同,证明重组菌株生成的产物为7α-羟基胆固醇。
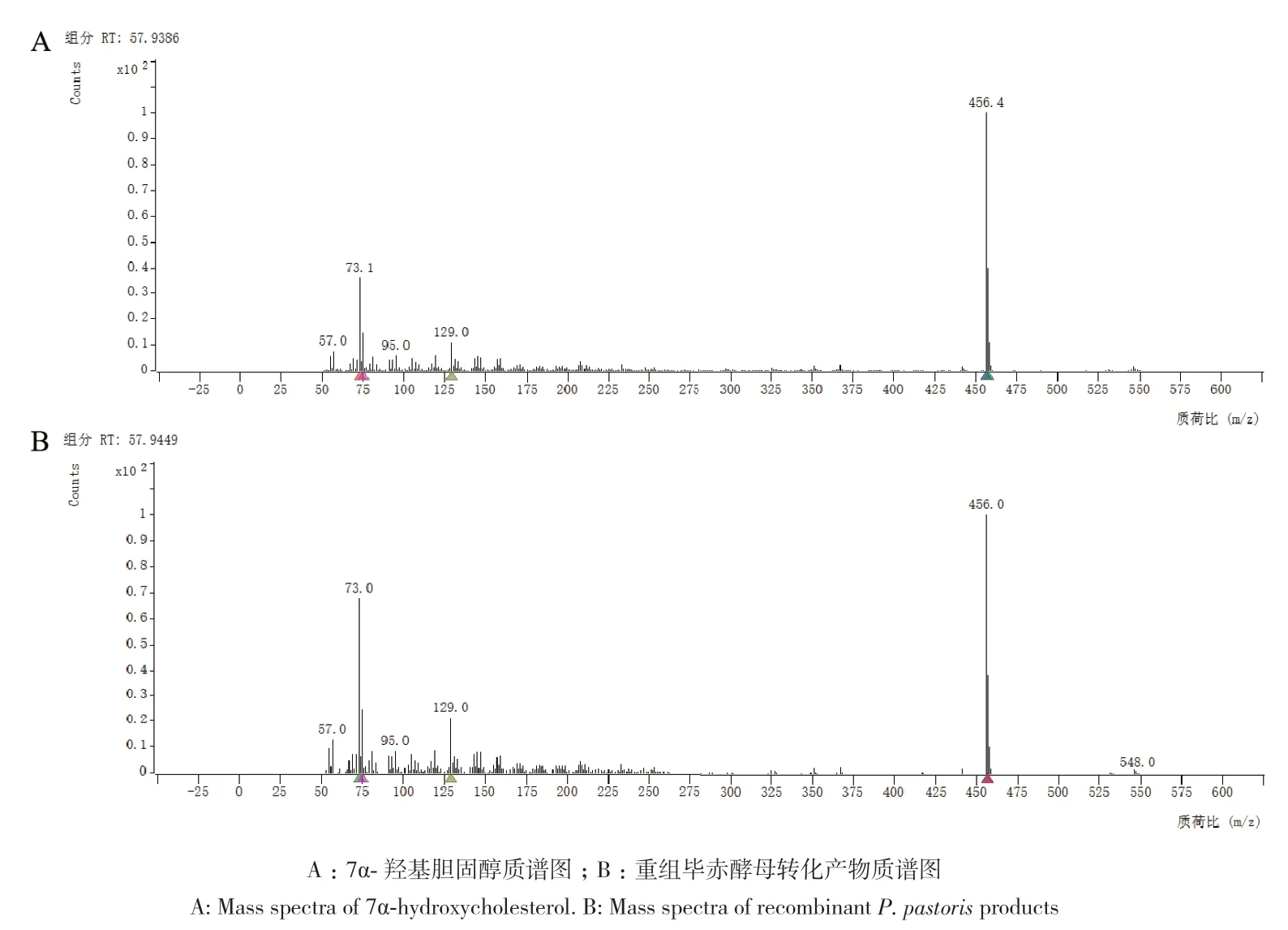
图3 重组毕赤酵母催化生成7α-羟基胆固醇的GC-MS分析图Fig. 3 GC-MS analysis of the 7α-hydroxycholesterol catalyzed by recombinant P. pastoris
2.2 CYP7A1的分子改造对7α-羟基胆固醇产率的影响
将利用分子生物学改造获得的5株突变株,进行催化胆固醇能力分析,转化产物经TLC分析(图2)、蒸发光检测(图4),与野生型相比发现,对111、114和284位点突变后酶的活性消失,说明这些位点对CYP7A1的活性有关键作用。对281和485的位点突变后,CYP7A1仍然具有活性,其中281位点的缬氨酸突变为丙氨酸后,活性稍有降低,为突变前的67.86%,而485位点的甘氨酸突变为丙氨酸后,活性提高,是野生型的108.71%,之后选取突变体G485A进行后续研究。
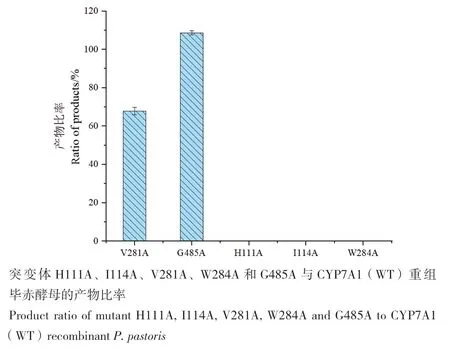
图4 突变毕赤酵母产率比较图Fig. 4 Production abilities of the mutants when compared with the wild type
2.3 不同来源CPR对重组毕赤酵母7α-羟基胆固醇产率的影响
将不同来源CPR与CYP7A1共表达,转化结果如图5所示,ScCPR、AnCPR、hCPR、RatCPR和SgCPR都能与CYP7A1适配,所获得的重组菌株催化胆固醇的能力均有提高,其中CYP7A1与SgCPR共表达使7α-羟基胆固醇的产率与CYP7A1野生型菌株相比提高了82.26%,但与人源CYP7A1同源的hCPR在共表达过程中适配性没有SgCPR显著。突变体G485A与SgCPR共表达毕赤酵母催化胆固醇的能力显著提高,与CYP7A1野生型菌株相比,产物产率提高了133.79%,产量为0.25 mg/L。
将全长的人源CYP7A1基因在毕赤酵母中进行表达,将毕赤酵母全细胞催化的发酵液进行萃取浓缩,用于薄层色谱(TLC)和气相色谱-质谱法(GC‑MS)分析,检测后确定发酵液中含有的产物为7α-羟基胆固醇。
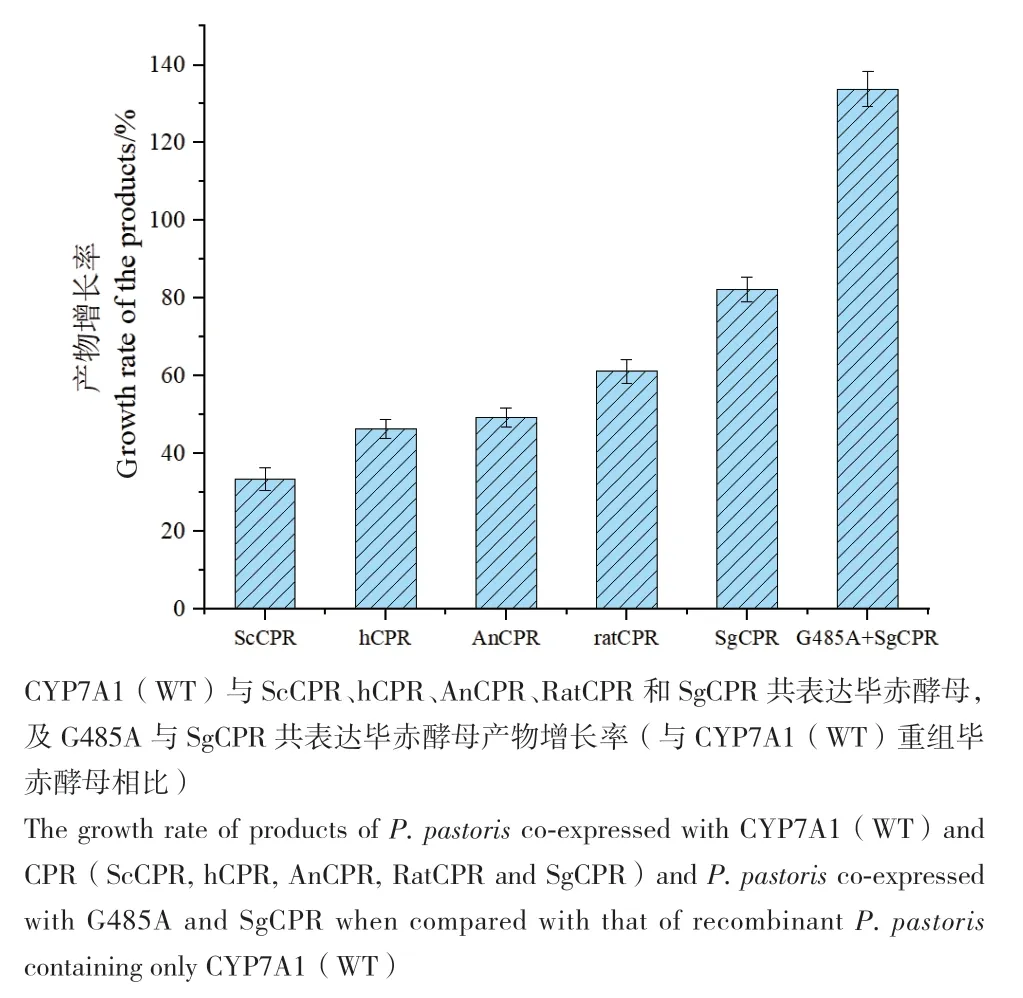
图5 CYP7A1与不同来源CPR共表达毕赤酵母对胆固醇转化能力的影响Fig. 5 Effects of P. pastoris co-expressed with CYP7A1 and CPR from different sources on cholesterol conversion
3 讨论
目前报道的胆固醇7α-羟化酶主要是在大肠杆菌中异源表达,但CYP7A1为膜蛋白,存在纯化步骤复杂、纯化蛋白浓度低、不能翻译修饰蛋白等问题。而且为了实现羟化酶在大肠杆菌中的高效表达,需要将N末端跨膜锚定结构域(氨基酸2-24)的氨基酸序列优化为MAKKT[14],并且为了在层析过程中溶解膜并保持蛋白质解聚,需要在纯化过程中添加表面活性剂,如Triton X‑100、Emulgen 913等[15]。不仅如此,还需要使用辛基氨基琼脂糖4B柱和阳离子交换层析等对蛋白进行进一步纯化[16]。为避免复杂的蛋白纯化步骤,实验选取毕赤酵母为表达宿主,它能识别真核基因,帮助其进行转录翻译后修饰,并且可以通过高密度发酵提高CYP7A1表达量和催化效率[17]。
3.1 突变体提高7α-羟基胆固醇产量的分析
目前对CYP7A1分子对接的研究主要集中在酶活性位点[18],而实验是通过Discovery studio软件的半柔性分子对接抓取与胆固醇小分子作用关键蛋白质位点,定点突变为丙氨酸。根据对接结果(图6),选取5个位点定点突变为丙氨酸,分别得到H111A、I114A、V281A、W284A和G485A突变体,这些位点存在于CYP7A1活性口袋附近,主要作用力为疏水作用力以及氢键相互作用,疏水作用可以维持活性中心空间结构的稳定性。对111、114及284位点突变后酶的活性消失,说明这些位点对CYP7A1的活性有关键作用,这些位点突变后可能改变了传递电子的能力或改变结合口袋的结构等。对281及485突变后催化活性变低或变高,可能是由于氨基酸极性不同导致的活性不同,氨基酸可分为亲水氨基酸和疏水氨基酸,亲水氨基酸对水分子具有一定的亲和性,疏水氨基酸对脂溶性物质的亲和性更高。由于胆固醇为非极性分子,根据“相似相溶”的性质,较容易与疏水氨基酸相互作用。实验中涉及的氨基酸极性从大到小的顺序为:His>Trp>Gly>Ala>Val>Ile,突变体485位点甘氨酸突变为极性更小的丙氨酸后,与胆固醇形成的疏水键的作用力更强,催化活性提高;突变体281位点缬氨酸突变为极性更大丙氨酸后,与胆固醇形成的疏水键的作用力减弱,催化活性降低;其次也可能是由于突变位点距离胆固醇C‑7位距离的不同导致活性不同,突变位点与胆固醇分子对接位置如图7所示。281位点与胆固醇C‑17位上的侧链相连,与C‑7位距离较远,而485位点与A环相连,与C‑7位距离较近,改变485位点氨基酸的大小对催化C‑7羟基化有较大的影响。综上所述281和485位点可以作为改造位点,将其突变为更加疏水的氨基酸或改变485位点氨基酸的大小会对酶与底物结合有进一步影响。
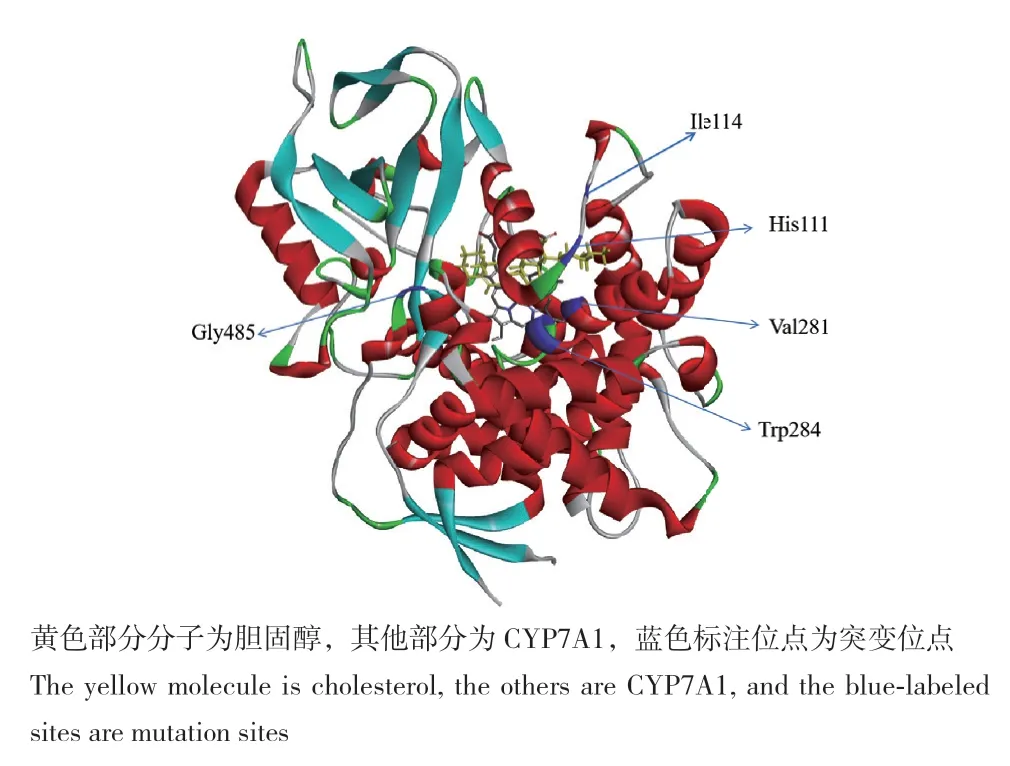
图6 CYP7A1与胆固醇分子对接结果图Fig. 6 Docking results of CYP7A1 and cholesterol molecules

图7 CYP7A1与胆固醇分子对接结果的二维平面图Fig. 7 2D floor plan of docking results of CYP7A1 and cholesterol molecules
3.2 CYP7A1(WT/突变型)与CPR适配提高7α-羟基胆固醇产量的分析
毕赤酵母为真核表达系统,自身含有CPR可帮助CYP7A1转移电子,但通过异源表达不同来源的CPR可增加产物7α-羟基胆固醇的产量。实验表明,ScCPR、AnCPR、hCPR、RatCPR和SgCPR与CYP7A1共表达菌株催化胆固醇的能力均有提高,并且SgCPR与G485A适配后催化能力更加显著。虽然CPR的过表达能够增强CYP7A1在毕赤酵母的羟基化水平,但并不是与CYP7A1基因同源的人源CPR与其适配性最好,可能是由于CYP‑CPR相互作用时,不同CPR中的FMN结构域附近特定残基组成不同导致的,FMN区域正电荷氨基酸数量显著影响其相互作用[19]。
既然幼儿园里吃到的水产品不足,那么家里就要每周至少吃一次水产品了。当然为了安全起见,应给孩子吃刺少的鱼比如龙利鱼、鳕鱼、桂鱼。这里特别要注意一类海鱼(如金枪鱼、沙丁鱼、三文鱼),青皮红肉,富含欧米伽3脂肪酸和虾青素,是适合全家人的健康食品,但是这类鱼同时富含组氨酸,如果加热时间不够长或者不新鲜都可能造成组胺中毒。所以,吃这类鱼一定要选新鲜的,然后充分加热。当然还可以吃虾,建议把虾去头之后再做,因为重金属都蓄积在虾头中。
因此在毕赤酵母中异源表达其他CYP450时,可添加外源CPR提高转化率。根据相关文献报道,CYP450与CPR相互作用中存在着一定的比例,选取合适的比例可以大大提高酶的催化活性[20],因此进一步实验可以通过CYP7A1适配不同浓度SgCPR,提高CYP7A1的转化能力。
3.3 重组毕赤酵母中生产7α-羟基胆固醇的分析
重组毕赤酵母生产7α-羟基胆固醇,是通过胆固醇7α-羟化酶在毕赤酵母异源表达的催化胆固醇实现的,其难点在于7α-羟基胆固醇的产量很低,只有产物浓度浓缩10-20倍才可用仪器检测到,因此并不是酶没有表达,而是产物浓度低到难以用蒸发光等仪器发现。并且胆固醇为脂溶性物质,难于进入细胞进行催化反应,即使在助溶剂的帮助下其溶解度也很低,因此找到胆固醇进入细胞的更有效办法,可显著提高7α-羟基胆固醇产量。
4 结论
构建CYP7A1异源表达平台并对其转化胆固醇能力进行研究,获得全细胞催化胆固醇生产7α-羟基胆固醇的重组毕赤酵母菌株,通过定点突变及适配不同来源CPR进一步提高了7α-羟基胆固醇产量。实验为7α-羟基胆固醇的高效生产奠定了良好的基础,也为CYP450的高效表达提供了有效方法。
