Loop B3对GH7内切纤维素酶功能的影响机制
2023-11-23杨俊钊张新蕊孙清扬郑菲
杨俊钊 张新蕊 孙清扬 郑菲
(北京林业大学生物科学与技术学院,北京 100083)
糖苷水解酶第七家族纤维素酶(glycoside Hydrolases family 7, GH7)在天然生物质转化和降解中具有重要作用,因此一直是许多研究的热点。在GH7中,主要包含外切纤维素酶和内切纤维素酶两大类,其中,内切纤维素酶全部来自真菌,作用于纤维素内部的非结晶区,随机水解分子内部的β-1,4糖苷键,将长链纤维素分子切割成短链[1];外切纤维素酶作用于纤维素链的还原/非还原性末端,将短链的纤维素分子切成纤维二糖[2]。目前,对GH7的研究主要集中在外切纤维素酶方面,而对内切纤维素酶的关注相对较少。CAZymes数据库(carbohydrate‑active enzymes, CAZymes)已被收录的具有性质报道的GH7内切纤维素酶仅有27个,大多在50-60℃和pH 4.0-5.0显示出最大酶活力[3-6]。根据PDB数据库(protein data bank, PDB)中收录的5个GH7内切纤维素酶晶体结构[ThCel7B(Trichoderma harzianum CBS 226.95, PDB ID: 5W0A)、TrCel7B(Trichoderma reesei, PDB ID :1EG1)、ReCel7B(Rasamsonia emersonii CBS 394.64, PDB ID: 6SU8)、FoCel7B(Fusarium oxysporum, PDB ID:1OVW)和HiCel7B(Humicola insolens, PDB ID:6YOZ)][7-11]可知,GH7内切纤维素酶的催化结构域是由6个loop区(A1、A2、A3、B1、B3和B4)围绕反向平行的β-折叠形成的β-三明治结构[12]。GH7内切纤维素酶中共有9个底物结合位点(‑7 -+2)[13]。其中,loop A1和B1位于底物结合位点‑7- ‑4处,loop A3和B3位于催化活性中心附近,loop B4位于产物出口处[11]。由于覆盖在催化口袋的loop较短,尤其是loop A3和loop B3的长度不能够完全包裹催化中心,使得活性中心更加暴露。
研究表明,催化口袋周围的loop参与酶的催化、底物结合过程,同时对酶的热稳定性也起着重要作用[14]。例如,GH12内切纤维素酶NfEG12A具有较长的loop 3结构,通过改变loop结构的长度,加强了底物和酶之间的氢键相互作用,从而提高了产物分子的转化率[15]。将GH5纤维素酶GtCel5 loop 6上的天冬酰胺Asn进行饱和突变后,筛选到N233G突变体的比活值及催化效率较野生型显著提高[16]。此外,loop结构对酶的热稳定性也起到重要作用,有研究人员将几丁质酶BcChi‑A的loop‑III截断后,其Tm值下降10℃[17]。GH7外切酶与内切酶的loop区具有类似的结构,在GH7外切纤维素酶中,loop结构参与酶的催化过程。研究人员将TrCel7A与纤维四糖、纤维戊糖和纤维己糖的复合物分别共结晶,观察到3个复合物结构中的loop B3区均参与底物的结合过程,且在催化过程中显示出较高的柔性[18]。研究人员通过截断TrCel7A loop B3中的8个残基(245GTYSDNRY252),验证得出酶对结晶纤维素的活性降低了50%,这表明loop B3是GH7酶催化的一个重要组成部分[19]。尽管有部分研究的关注点在GH7外切酶的loop区上,但对于内切纤维素酶loop区的功能仍缺乏相关研究。经分析,在GH7内切纤维素酶loop区中,loop B3位于催化中心附近[11],其氨基酸序列长度可分为两种类型:一类是包含18个氨基酸残基的长链型;另一类是包含4个氨基酸的短链型。据研究显示,大多数耐热性能比较好的GH7内切纤维素酶均包含短链型的loop B3结构[3-5,11]。为了探究loop B3在GH7内切纤维素酶中的影响效果及其机制,本文以嗜热毁丝菌M.thermophila来源的GH7纤维素酶MtCel7b为核心研究材料,通过截短其长链loop B3中的14个氨基酸残基(216GPYLCEGAECEFDG229),构建B3cut突变体,并分析比较突变体与野生型之间在酶催化效率及热稳定性方面的差异,为GH7内切纤维素酶loop区的功能研究提供理论支持。
1 材料与方法
1.1 材料
1.1.1 基因和载体 Mtcel7b 基因编码序列由北京睿博兴科生物技术有限公司合成;克隆宿主大肠杆菌感受态细胞Escherichia coli Trans1‑T1购自北京全式金生物技术有限公司,用于目的基因的克隆。表达质粒pPIC9和表达宿主毕赤酵母Pichia pastoris GS115购自美国Invitrogen生命技术有限公司,用于表达载体的构建和外源基因的表达。
1.1.2 主要试剂与仪器 无缝克隆与重组试剂盒、快速凝胶提取试剂盒购自北京全式金生物技术有限公司;质粒提取试剂盒购自天根生化科技(北京)有限公司;胰蛋白胨、酵母浸粉购自赛默飞世尔科技公司;底物羧甲基纤维素钠(CMC‑Na)、地衣多糖、大麦葡聚糖购自Sigma‑Aldrich。琼脂糖购自Biowest公司、YNB购自北京兰博利德生物技术有限公司。D‑无水葡萄糖和琼脂粉均购自博奥拓科技有限公司。
PCR仪,Bio‑Rad T100型梯度PCR仪分子扩增仪;蛋白电泳仪,DYY‑8C电脑三恒多用电泳仪;培养箱,一恒Blue pard经济型生化培养箱;分子过滤膜包,赛多利斯回旋流切向流超滤器超滤膜包VF20P4;酶标仪,瑞士Tecan M200 PRO多功能酶标仪;蛋白纯化仪,ÄKTA Pure;核酸蛋白测定仪,赛默飞NanoDrop2000超微量分光光度计。
1.2 方法
1.2.1 序列与结构分析 使用ExPASy‑ProtParam tool(https://web.expasy.org/protparam/)对蛋白的理论分子量和等电点进行分析,使用SignalP 5.0(http://www.cbs.dtu.dk/servi Ces/Signa LP/)对蛋白的信号肽进行预测,利用BLASTp(Basic Local Alignment Search Tool protein)对蛋白序列进行同源性分析,利用Smart(http://smart.embl‑heidelberg.de/)对蛋白质的结构域进行预测,利用AlphaFold2对蛋白质建模。
1.2.2 质粒构建与提取 将目的基因Mtcel7b重组至pPIC9表达质粒EcoR I/Not I克隆位点处,利用热激法转化至Escherichia coli Trans1‑T1细胞中,后涂布于固体LB培养基[0.5%酵母浸粉(W/V),1%胰蛋白胨(W/V),1%氯化钠(W/V),1.5%琼脂粉(W/V)]上,37℃倒置培养。12 h后挑取单克隆进行菌落PCR验证并送测序,后将测序正确的克隆子接入到液体LB培养基[0.5%酵母浸粉(W/V),1%胰蛋白胨(W/V),1%氯化钠(W/V)]中,在37℃条件下培养12 h,然后提取质粒。突变体B3cut的构建是以Mtcel7b为模板,以MtCel7b‑F1′(5′‑GA GAGGCTGAAGCTTACGTAGAATTC‑3′),MtCel7b‑R1′(5′‑AGGCGAATTAATTCGCGGCCGC‑3′);B3cut‑F2′(5′‑CTCATCCTTGTTCCAAGGCTGTATGTGATAAGAA TGGTTGCGCCTG‑3′),B3cut‑R2′(5′‑CAACCATTCTTA TCACATACAGCCTTGGAACAAGGATGAGGAG‑3′)为两组特异性引物进行PCR扩增,利用overlap PCR方法获得突变体基因的全长。PCR反应程序为:95℃预变性3 min;95℃变性30 s,55℃退火30 s,72℃延伸30 s,30个循环;72℃终延伸5 min;4℃保存。质粒提取方法同Mtcel7b。
1.2.3 蛋白的表达与纯化 重组质粒pPIC9‑Mtcel7b用限制性内切酶Dra I线性化处理,经核酸电泳纯化后回收线性化产物,通过电击转化至毕赤酵母GS115感受态细胞,然后涂布在固体MD培养基[1.5%琼脂糖(W/V),2%葡萄糖(W/V),灭菌后加入1.34% YNB(W/V),4×10-4生物素(W/V)]上。在30℃倒置培养48 h后,筛选出重组转化子。用牙签随机挑选48个克隆子,在固体MD平板上点种,并在30℃恒温箱内倒置培养48 h。同时,将牙签挑取的克隆子接种于含有3 mL BMGY培养基[1%酵母浸粉(W/V),2%胰蛋白胨(W/V),1%甘油(W/V),灭菌后加入1.34% YNB,4×10-4生物素(W/V)]的酵母管中。在30℃,200 r/min培养48 h后离心去除上清液,菌体用1 mL BMMY培养基[1%酵母浸粉(W/V),2%胰蛋白胨(W/V),灭菌后加入1.34% YNB,4×10-4生物素,0.5%甲醇(V/V)]重悬,诱导培养48 h后离心收集上清液。利用3,5‑二硝基水杨酸(DNS)法测定酶活,并用SDS‑PAGE检测是否表达出目的蛋白。将筛选出的高活性阳性克隆子接种至30 mL YPD液体培养基[1%酵母浸粉(W/V),2%胰蛋白胨(W/V),2%葡萄糖(W/V)]中,30℃,200 r/min培养48 h后,以 0.5%(V/V)的接种量接种至BMGY液体培养基中。30℃,200 r/min培养48 h后,收集菌体转移至BMMY诱导培养基中,30℃,200 r/min继续诱导培养48 h,每隔24 h补加一次甲醇[0.5%(V/V)]。48 h后收集上清液,利用10 kD膜包浓缩至15 mL体积后进行脱盐处理,脱盐处理后的酶液加入到平衡后的HiTrap QXL阴离子交换层析柱中,在10 mmol/L的pH 8.0的Tris‑HCl缓冲液中,使用0-1 mol/L NaCl进行线性梯度洗脱,收集相应的峰所对应的蛋白并进行酶活性的测定和SDS‑PAGE分析。
1.2.4 酶活力的测定 纤维素酶的酶活性测定采用DNS法,具体反应体系为50 μL适当稀释的酶液加入到450 μL底物中,恒温中反应10 min,后加入750 μL DNS 终止反应,沸水煮5 min后立即置于冷水中冷却至室温。从反应体系中吸取250 μL在540 nm下测定吸光值,根据标准曲线计算出相应酶活值,每组实验设置3个重复。酶活定义为:在酶的最适条件下,每分钟产生1 μmol还原糖所需的酶量为1个单位(U)。
2014年11月10日晚,参加北京APEC会议的各国领导人携配偶在欢迎晚宴前现身,身着特色中式服装合影的他们,吸引了全世界的目光。国家主席习近平说:“看到各位都穿上中国式服装,既充满中国传统元素,又体现了现代气息,让我们更感亲近,中国老百姓看了以后也会感到亲切,就像到邻居家串门。尤其是女装,女士穿上更显秀美!”
1.2.5 酶学性质分析
1.2.5.1 最适温度和温度稳定性的测定 将纯化后的酶进行适当稀释,选择10 mg/mL CMC‑Na作为底物,温度范围设定为40-70℃,重组酶在最适pH条件下反应10 min,测定酶活。将最高酶活性设置为100%,并计算其他温度下的活性作为相对值(%),并绘制最佳温度趋势图。将重组酶分别在70、80和90℃下分别孵育2、5、10、30和60 min后,在最适条件下测定酶活力。以未处理酶液的酶活值作为100%,计算各时间点和温度点下的相对酶活值并绘制温度稳定性趋势图。
1.2.5.2 最适pH及pH稳定性的测定 重组酶和底物(10 mg/mL CMC‑Na)分别用不同pH缓冲液(pH 3.0-7.0:100 mmol/L 柠檬酸-磷酸氢二钠;pH 8.0-9.0:100 mmol/L Tris‑盐酸;pH 10.0-12.0:100 mmol/L 甘氨酸-氢氧化钠)进行稀释,将重组酶放置在最适温度条件下反应10 min,测定各pH条件下的剩余酶活。以酶活最高的pH条件下的酶活值作为100%绘制最适pH和pH稳定性趋势图。
1.2.5.3 比活值测定 以450 μL的10 mg/mL CMC‑Na、10 mg/mL大麦葡聚糖、10 mg/mL地衣多糖为底物,分别向其中加入50 μL适当稀释的酶液,在重组酶最适条件下反应10 min,测定重组酶的比活值。
1.2.5.4 动力学常数的测定 以浓度范围为4-20 mg/mL的CMC‑Na为底物,反应体系包含450 μL的底物和50 μL的酶液,在最适条件下反应5 min,测定酶活性。利用双倒数法作图并计算得到Km、Vmax、kcat及kcat/Km的值。
1.2.5.5 耐盐性和盐稳定性的测定 重组酶耐盐性的测定是将CMC‑Na溶于不同浓度的NaCl(1-4 mol/L)溶液中,在最适条件下,与适当稀释的酶液孵育10 min,测定剩余酶活力。在测定酶的盐稳定性时,将重组酶与不同浓度的NaCl(1-5 mol/L)混合后在37℃下共同孵育1 h,后取50 μL孵育后的酶液与450 μL CMC‑Na在最适条件下反应10 min。以酶活最高的盐浓度条件下的比活值为100%绘制耐盐性和盐稳定性趋势图。
2 结果
2.1 序列与结构分析
来源于M. thermophila的GH7内切纤维素酶编码基因Mtcel7b(GenBank Accession No.:XP_003664606.1)核苷酸序列全长为1 368 bp,编码456个氨基酸,前20个氨基酸为信号肽序列,其去除信号肽后的理论分子量为47.2 kD,等电点为4.75。MtCel7b的结构预测模型表明(图1‑A),MtCel7b的结构与其他GH7家族纤维素酶一样,由6个loop区围绕反向平行的β-折叠形成的β-三明治结构。多序列比对结果(图1‑B)显示,E194、D196、E199为催化三联体,其中,E194和E199分别作为亲核试剂和酸/碱试剂催化糖苷键断裂(编号不包括信号肽序列)。MtCel7b的loop B3比其他短链的内切纤维素酶多出14个氨基酸残基。MtCel7b长链的loop B3额外形成了一个β-折叠片和一个α-螺旋结构。
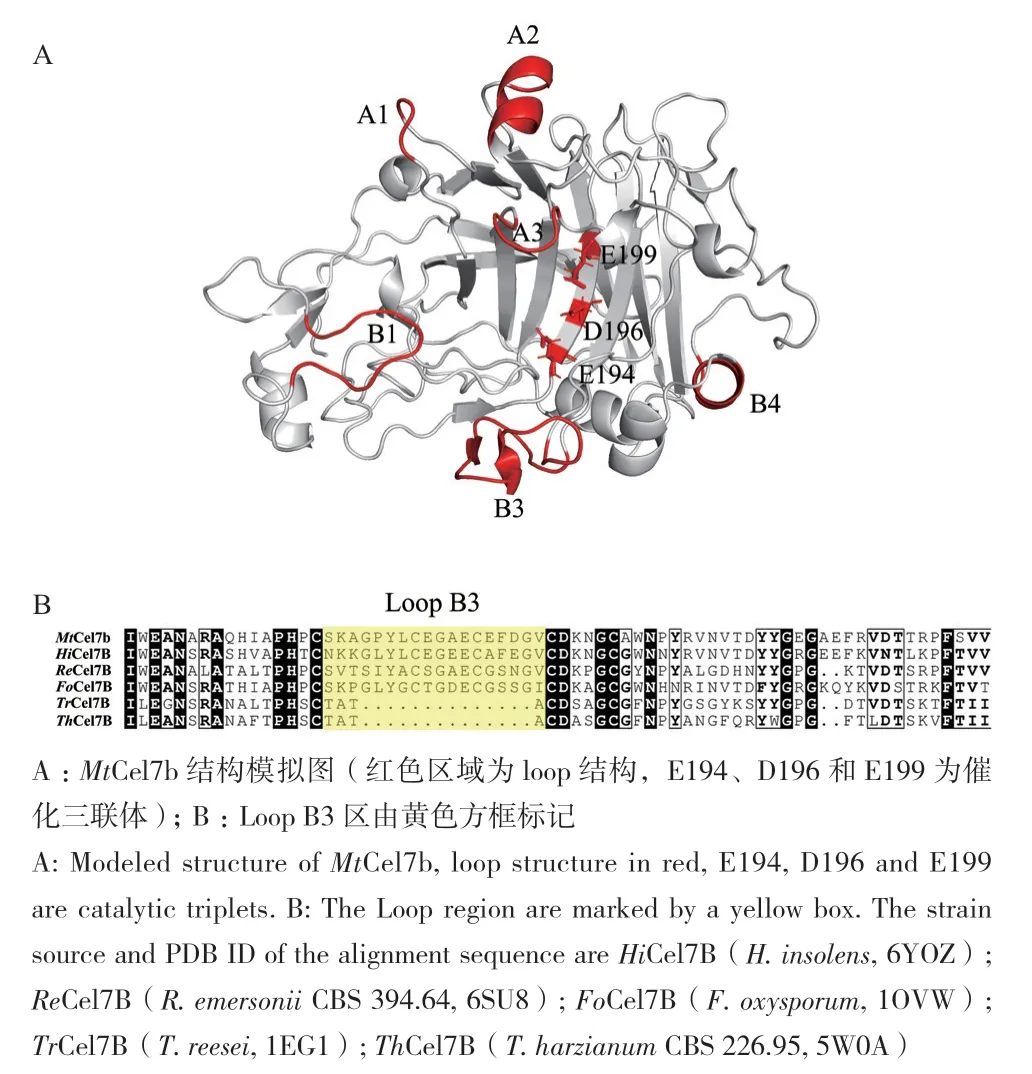
图1 MtCel7b结构模拟(A)及氨基酸序列对比(B)Fig. 1 Structural simulation(A)and amino acid sequence alignment(B)of MtCel7b
2.2 MtCel7b及其突变体的表达和纯化
SDS‑PAGE结果显示(图2),野生型MtCel7b与突变体B3cut的表观分子量均大于其理论分子量(MtCel7b: 47.2 kD;B3cut: 45.7 kD)。经预测,野生型与突变体均包含3个N糖基化位点,可能是造成表观分子量大于理论分子量的原因之一。
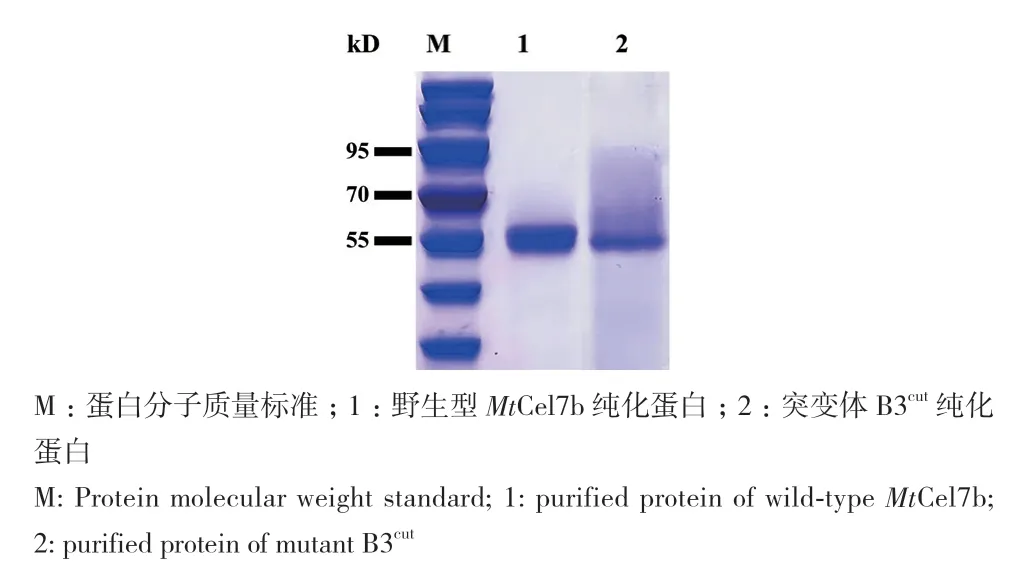
图2 野生型MtCel7b和突变体B3cut的SDS-PAGE分析Fig. 2 SDS-PAGE analysis of wild-type MtCel7b and mutant B3cut
2.3 MtCel7b及其突变体的酶学性质分析
2.3.1 最适温度及温度稳定性分析 在pH 5.0条件下测定野生型及突变体在不同温度(40-70℃)下的酶活力,由图3‑A可知,MtCel7b的最适温度为60℃,在40-60℃之间的酶活力均在80%以上。将MtCel7b分别在70、80和90℃下孵育,检测其剩余酶活力(图3‑D-F)。结果表明,MtCel7b具有良好的耐热性,在80℃和90℃条件下处理5 min后,剩余酶活仍保持在80%以上(图3‑E, F)。此外,MtCel7b在90℃孵育1 h后仍可检测到40%以上的酶活性。突变体B3cut的最适温度为50℃,整体趋势较野生型变窄(图3‑A)。突变体在70、80和90℃的热稳定性较野生型提高,尤其在90℃孵育1 h后,其剩余酶活高达56%(图3‑F)。
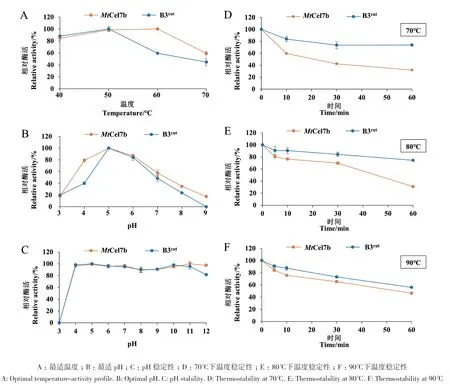
图3 野生型MtCel7b及突变体B3cut的酶学性质Fig. 3 Enzymatic properties of wild-type MtCel7b and mutant B3cut
2.3.2 最适pH及pH稳定性分析 在pH 3.0-9.0范围内测定最适pH值(图3‑B)。MtCel7b的最适pH为5.0,在pH 5.0-6.0范围内保留大约80%以上的酶活性。B3cut与野生型一致均在pH 5.0下表现出最佳酶活性,但突变体的pH耐受范围缩小(图3‑B)。在pH 4.0时,野生型的相对酶活性高达79%,而突变体的相对酶活性为40%。此外,MtCel7b和突变体B3cut在pH 4.0-12.0稳定性较好,在37℃下处理1 h后,仍然具有90%以上的酶活(图3‑C)。
2.3.3 耐盐性和盐稳定性分析 在0-4 mol/L和0-5 mol/L NaCl中,测定了耐盐性和盐稳定性。在耐盐性方面,MtCel7b的相对比活随着底物中盐浓度的增加而逐渐降低(图4‑A),即当2 mol/L时,MtCel7b的相对比活值为55%,而当底物中盐浓度增加至4 mol/L时,MtCel7b的相对比活值下降至37%。盐稳定性检测中,当酶液分别与0-5 mol/L NaCl缓冲液孵育1 h后,其野生型的比活值保持恒定,具有稳定的耐盐性(图4‑B)。进一步分析显示,虽然突变体B3cut经过0-5 mol/L NaCl溶液分别处理后的活性变化趋势与野生型一致,但是其在不同浓度下的相对比活值明显高于野生型,说明突变体B3cut在0-5 mol/L NaCl的浓度下稳定性更好。

图4 野生型MtCel7b及突变体B3cut的耐盐性(A)与盐稳定性(B)Fig. 4 Halo tolerant(A)and halo stability(B)of wild-type MtCel7b and mutant B3cut
2.3.4 比活性和动力学分析 以10 mg/mL CMC‑Na、大麦葡聚糖和地衣多糖为底物测定MtCel7b及其突变体的比活性。MtCel7b水解CMC‑Na、大麦葡聚糖和地衣多糖的比活值分别为(38 ± 2)、(790 ± 1)和(493 ± 10)U/mg(图5)。突变体B3cut表现出与野生型相似的底物偏好,但B3cut水解这3种底物的比活值分别降低至(25 ± 1)、(255 ± 1)和(130± 3)mg/mL,较野生型降低了34%-74%。
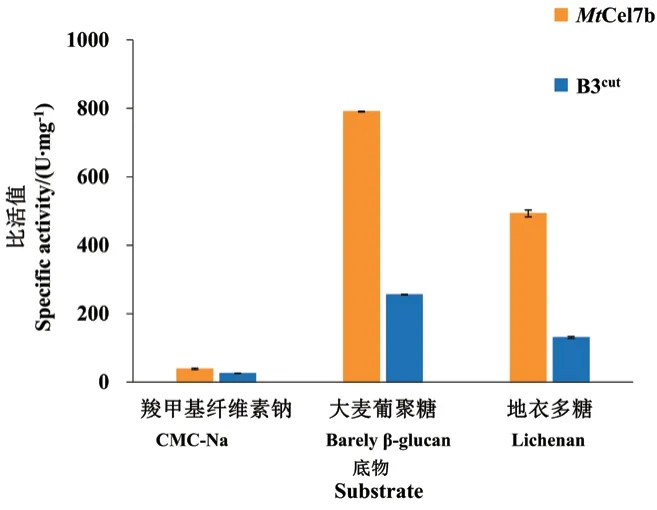
图5 野生型MtCel7b及突变体B3cut的比活值Fig. 5 Specific viabilities of wild-type MtCel7b and mutant B3cut
以4-20 mg/mL CMC‑Na作为底物,在最适反应条件下测定酶的动力学参数。MtCel7b的Km和Vmax值分别为(8.8 ± 0.8)mg/mL和(143± 7)μmol/(min·mg),kcat/Km为(12.8 ± 0.1)mL/(mg·s)。与野生型相比,突变体的Vmax[(69± 8)μmol/(min·mg)]较野生型降低52%,催化效率[(4.9 ± 0.6)mL/(mg·s)]比野生型低约62%,并且突变体对底物的亲和力减弱[Km=(10.9± 3.3)mg/mL]。
2.4 分子动力学模拟分析
根据图6的RMSD的结果来看,MtCel7b与突变体在50 ns后达到平衡,平均RMSD值均为2.0 Å。MtCel7b与突变体均在loop区域表现出相似的RMSF趋势,波动越大,显示出loop区的灵活性越强。突变体中与loop B3相邻的257-266位残基的RMSF值增加到5 Å,而野生型相应位置的RMSF值仅为2 Å(蓝色虚线框)。说明截短loop B3后,会影响相邻结构的波动情况。
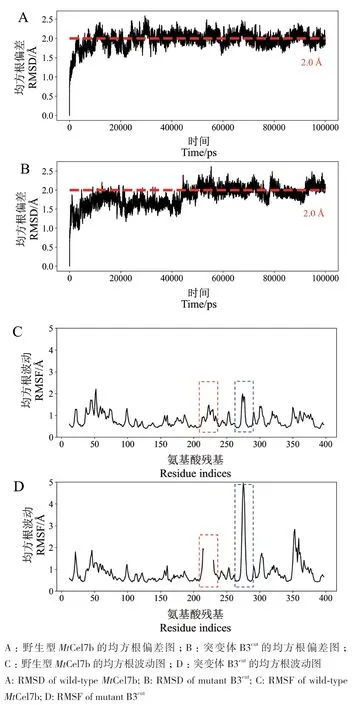
图6 野生型MtCel7b及突变体B3cut的均方根偏差和均方根波动图Fig. 6 RMSD and RMSF of wild-type MtCel7b and mutant B3cut
图7‑A, B的主成分分析图说明野生型与突变体均具有5种不同的状态,分别以不同颜色显示。野生型的样点分布分散,表明每种状态之间的相似性较低,结构变化较大。但是,突变体状态内与状态之间的聚类情况良好,样点分布更加集中,状态间有较好的区分,说明其在100 ns分子动力学模拟过程中构象相对稳定。提取分子动力学模拟的平均结构用于进一步分析,根据预测蛋白活性口袋大小的在线网站POCASA(http://altair.sci.hokudai.ac.jp/g6/service/pocasa/)计算得到野生型MtCel7b和突变体B3cut活性口袋的体积分别为2 799 Å3和1 078 Å3,野生型的口袋体积大概是突变体的2.5倍(图7‑C,D)。图7‑E, F显示,B3cut的loop A3(351GDN353)和loop B1(W52)均向催化活性中心移动,导致其催化通道较野生型更加密闭。从平均结构看出,野生型的loop B3周围有一个复杂的氢键相互作用网络(图7‑E)。C225主链羧基O和主链氨基H分别与K233侧链氨基HZ2和G222主链羰基O形成侧链-主链氢键和主链氢键,并且C225还与C220的巯基形成二硫桥。在复杂的氢键网络的控制下,K233侧链氨基HZ3和主链氨基H分别与E226主链羰基O和C231主链羰基O形成侧链 - 主链氢键和主链氢键。然而,由于loop B3截短,K233失去周围氢键束缚,导致其侧链扭转了120°,并且侧链氨基HZ2与催化残基E194的侧链羧基OE1形成盐桥,并且K233的主链氨基H和D232侧链羧基OD2形成新的侧链-主链氢键,使两个催化残基(E194、E199)的Cα距离从12.7 Å减少到11.7 Å,并且明显缩小了催化口袋。
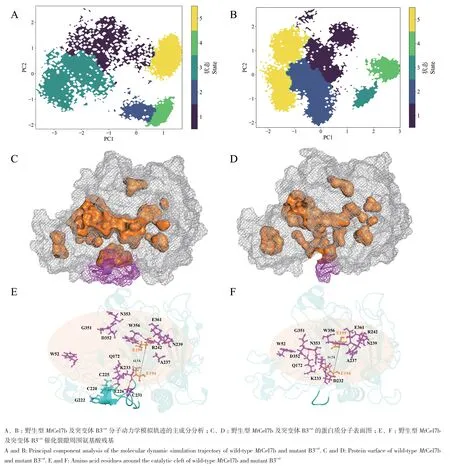
图7 野生型MtCel7b及突变体B3cut分子动力学模拟的轨迹主成分分析及平均结构图Fig. 7 Trajectory principal component analysis plots and average structure of molecular dynamics simulations of wild-type MtCel7b and mutant B3cut
除此之外,催化通道两端残基的位置也发生了改变。图7‑C, D表明,突变体的催化口袋明显较野生型窄。如图所示,残基W52、Q172、K233、N234、R242、G351、D352、N353、W356和E361均向催化中心移动。在催化通道的‑3位处,突变体B3cut中的loop B1中W52侧链的方向发生了变化,与野生型相比更接近催化中心。B3cut的loop A3中D352主链羰基O和靠近loop B3的Q172侧链酰基2HE2之间形成侧链-主链氢键,导致催化口袋被周围的氨基酸覆盖,催化通道的入口变窄。同时,在催化通道的+2位点处,氨基酸的取向也发生了变化,从而引起了氢键的改变。在野生型中,R242侧链胍基2HH2和主链氨基H只与W356主链羰基O和N239侧链酰基OD1之间形成侧链-主链氢键(图7‑E),而在B3cut中,R242周围形成了一个新的复杂的氢键网络。如图7‑F所示,R242主链氨基H、侧链胍基1HH1、2HH2、HE分别与催化裂隙上方的N239侧链酰基OD1、A237主链羰基O和裂隙下方的E361侧链羧基OE1、OE2之间形成侧链-主链氢键,这使得催化通道正位处变窄。虽然R242没有与W356形成氢键作用力,但W356的吲哚环旋转了50°,使产物生成部位更加封闭,增加了催化裂隙的疏水性。
3 讨论
MtCel7b的最适温度和最适pH值与大多数真菌源的GH7内切葡聚糖酶相似,均在50-60℃和pH 4.0-5.0的范围内显示出最大的酶活性[3-6]。此外,与先前的报道一致,MtCel7b对大麦葡聚糖的比活值高于对CMC‑Na的水解活性,这可能是由于CMC‑Na的分子结构被甲氧基侧链高度取代,从而干扰酶的活性[20]。目前报道的大多数GH7内切酶都属于嗜中温酶,例如,来源于Aspergillus oryzae KBN616的CelA和CelB分别在60℃和55℃的温度处理10 min后迅速失活[3],当在60℃条件下处理10 min后,来自H. grisea var. thermoidea的EGI稳定性瞬间下降[4]。来源于A. fumigatus的Af‑EGL7在60℃的温度下处理30 min基本失活[6]。相比之下,MtCel7b的耐热性显著优于其他已知的GH7内切纤维素酶,在90℃的温度下处理1 h后仍能剩余40%以上的酶活,这大大提高了MtCel7b在工业应用上的潜力。除此之外,MtCel7b也表现出良好的耐盐性。有研究表明,嗜盐蛋白的标志之一是表面具有较高的负电位,蛋白质表面过量的酸性氨基酸残基有助于增强水分子和金属离子的结合,有助于形成水化膜,保护并维持蛋白质的结构完整性和活性[21-22]。在本研究中,MtCel7b蛋白表面具有大量负电荷残基。其中,MtCel7b有64个带负电荷的残基,天冬氨酸占总氨基酸残基的7.1%,谷氨酸占7.6%。B3cut有60个带负电荷的残基,天冬氨酸和谷氨酸均占总氨基酸残基的7.1%。蛋白质表面大量的酸性残基也使得酶具有较低的pI(pI MtCel7b= pIB3cut= 4.86)。纤维素酶作为工业应用的关键酶系,需要适应工业操作中特殊的物理和化学环境[23]。例如,在食品加工过程中,会遇到高温(如烘焙、烘干等)、低温(如低温发酵、果汁澄清等)、高压(如研磨、挤压等)、高渗(如盐渍、糖渍等)、强酸、强碱等较为苛刻的环境条件[24]。所以,能适应极端环境的酶备受工业青睐。MtCel7b在酶学性质上展现出的良好的耐热、耐盐性,表明其具有一定的潜在研究价值和工业应用价值。Loop区结构及氨基酸组成是影响酶热稳定性及活性的一个主要因素,尤其是催化通道周围的loop区[25]。本研究中,将MtCel7b loop B3截短后发现,突变体B3cut虽然活性降低了,但其在高温下的热稳定性较野生型有所提高。分子动力学模拟结果表明,loop B3的局部变化使酶的整体构象发生了较大程度的改变,特别是底物通道两端,结构更加紧凑,从而在高温环境下保持稳定。此外,loop B3的缺失也使催化通道内的氢键作用网络重新排布,尤其是催化残基周围的区域。在突变体中,K233位点丧失了与loop B3之间的联系,与催化残基E194建立了新的盐桥,导致两个催化位点之间的空间距离改变,从而使突变体的活性显著下降。Km值的增大也从另一个角度说明,突变体催化通道内氢键网络的变化使酶结合底物的能力有所减弱。综上所述,野生型loop B3中氨基酸之间的氢键相互作用网络的影响使得MtCel7b具有更开放的催化裂隙,可以招募更多的底物分子,而突变体由于催化裂隙的开口变小,可能会干扰酶对底物的抓取和释放,因此,突变体的催化效率较野生型显著降低。本研究丰富了GH7嗜热内切纤维素酶的酶资源库,同时为纤维素酶loop结构的研究提供了理论支持。
4 结论
本研究从M. thermophila克隆表达了一个新型嗜热、盐耐受度高的GH7内切纤维素酶MtCel7b,通过构建loop B3截短突变体,探究了loop B3对GH7内切纤维素酶热稳定性及催化活性的重要影响。酶学性质与分子动力学模拟结果显示,MtCel7b中长链的loop B3可以增加催化活性通道的体积,使其在催化底物过程中更加灵活。截短loop B3后,导致突变体B3cut催化通道体积的减小,酶的催化效率降低,热稳定性增强。
