生鲜肉持水性机理研究进展
2023-04-06余青青高瑞昌包玉龙
余青青,刘 娇,洪 惠,高瑞昌,包玉龙,*
(1.江苏大学食品与生物工程学院,江苏 镇江 212013;2.中南民族大学生命科学学院,湖北 武汉 430074;3.中国农业大学食品科学与营养工程学院,北京 100083)
新鲜瘦肉大约含有75%的水分,持水性是肉品在外力作用下,保持自身水分或后续加工中外加水分的能力。贮藏加工过程中,肉中水分处在动态变化之中,水分可以因蒸发、渗出和烹饪等过程损失,也可以在氯化钠、多聚磷酸盐、酸碱腌制液作用下而增加[1]。持水性是生鲜肉最重要的品质之一,肉品持水性不仅与感官品质和营养价值相关,也影响肉品的贮藏特性。质地、多汁性、颜色、滋味、营养素的保留以及微生物的生长均与肉品中水分含量及水分结合能力有关[2]。因此,肌肉的持水性能一直是肉品行业关注的重点。
有关肉品持水性机理的研究开展较早,20世纪60年代,Hamm[3]提出以肌原纤维溶胀为核心的肉品持水性机制,认为静电相互作用是肉品持水的基础;此后Offer等[1]从肌肉结构的角度进一步发展了Hamm提出的肌肉持水理论,指出渗透压的形成对于肌肉水分的保持有重要作用;Tornberg[4]认为除上述因素外,肌肉内部存在许多毛细结构,而毛细管作用力可以截留大量的水分。生鲜肉宰后成熟过程中,僵直横桥的形成以及内源酶导致的蛋白降解对于持水性有重要的调控作用。Kristensen等[5]研究发现,宰后成熟过程中,猪肉的持水性先下降后上升,由此提出细胞骨架蛋白在宰后成熟过程中的降解可以导致肌纤维外汁液回吸到肌纤维内部的猜想;Huff-Lonergan等[6]系统阐述了宰后僵直过程中肌纤维间和肌束间空隙对生鲜肉汁液流失的影响。随着相关学科的发展,持水性的理论也在不断更新,如Puolanne等[7]引入了低密度水、高密度水等全新的概念,对肉品持水性的理论基础进行了深入探讨。近年来,蛋白质氧化等化学修饰对于肉品持水性的影响也引起了广泛的研究兴趣[8-10]。本文首先阐述肌肉微观结构和其中水分分布,接着介绍肉制品持水的理论基础,并简介常见的持水性测定和表征方法,最后详细讨论影响肉制品持水的理化因素及其可能的机制,以期为肉类研究提供理论基础,为肉类产品的加工提供参考。
1 肌肉微观结构及水分分布
肌肉组织的微观结构高度有序,按照从宏观到微观的层级,可依次分为肌束、肌纤维、肌原纤维、肌丝(图1A)。一般而言,水分被紧密束缚在肌肉各个结构单元内,约占新鲜肌肉质量的75%,其中约85%的水分位于肌纤维内部,被束缚在由粗细肌丝组成的稠密的肌原纤维网络之中,而另外约15%的水分则位于肌纤维外部。肌肉中的水分空间分布位置包括:肌原纤维内与肌原纤维之间、肌原纤维和肌纤维膜之间、肌纤维之间、肌束之间。肌肉中的水分根据流动性可分为结合水、不易流动水、自由水(图1A)。场核磁共振技术通过检测氢质子弛豫时间证明了这3 种状态水分的分布[11-13]。结合水由于与蛋白质表面有很强的相互作用而拥有非常稳定的性质,只有在煅烧的情况下才会解离,因此不易受到外界因素,如剪切力、重力、加热或冷冻等的影响。结合水仅占为总水分含量的很小一部分,大约每克肌肉蛋白中含0.5 g结合水[14],相当于鲜肉质量的8%左右。也有学者对此持不同观点,认为肌肉中只有大约0.5%的水分是真正的结合水[15]。结合水之外的水分根据所处位置分为不易流动水和自由水,不易流动水又称截留水,位于肌原纤维之内,而自由水主要分布在肌原纤维之间以及肌纤维外部[16]。肌肉在宰后成熟过程中,横桥连接的形成、骨架蛋白的降解、蛋白质变性等原因导致微观结构变化,从而改变细胞内外空间形成的汁液流失通道,进而影响持水性[5,17-18]。
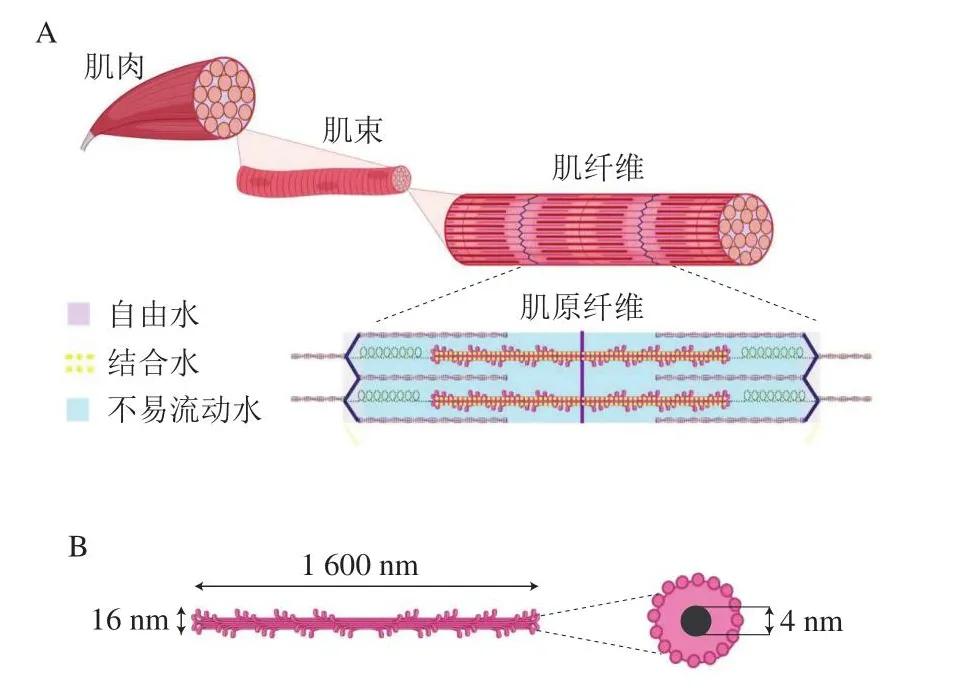
图1 肌肉微观结构及水分分布示意图[7,16]Fig.1 Illustration of the microstructure of muscle and the distribution of water in muscle matrix[7,16]
尽管已有大量实验研究了肌肉微观结构及其对肉类品质的影响[19-20],但在更细微尺度上,如粗细肌丝结构与不易流动水间的关系仍然需要进一步的探明。粗肌丝(以肌球蛋白为主)大约含有200~300 个肌球蛋白分子,构成了约16 nm×1 600 nm的纤维[21]。有学者指出,粗肌丝中心含一个直径约4 nm的中空部分[22](图1B),虽然这一结构尚未得到证实,但粗肌丝中心的中空部分很可能与肉品持水性有很大关联。随着超分辨显微镜等技术的出现[23],肌肉微观结构有望进一步探明。
随着对于肌肉结构的认知不断加深,科研人员对肌肉内水分的理化性质也有了新的发现,有助于更好地理解肉品的持水性。对于肌纤维内的水分,有学者认为由于排布高度结构化,水分的渗透压和流动性都与普通的体相水不同,有时甚至表现为一种类玻璃态[24]。离子和其他一些物质根据其与水分子相互作用的特点,可以分为亲液型(水结构增强)和离液型(水结构破坏)。亲液基团(包括多价离子和半径较小的离子)诱导水分子排列成有序结构,形成类似玻璃态的状态,称为低密度水。相反地,离液基团(大的单价离子)诱导水分子呈无序结构,形成高密度水(图2)。Puolanne等[7]猜测高密度水具有更低的黏度和更好的流动性,与临近肌肉结构的结合较为松散,因而其含量越高则持水性越低。已有研究表明肌肉中存在高密度水,如肌动蛋白丝附近的水化层[25]。但目前关于高密度水/低密度水对肉品持水性的影响还有待进一步的阐明。
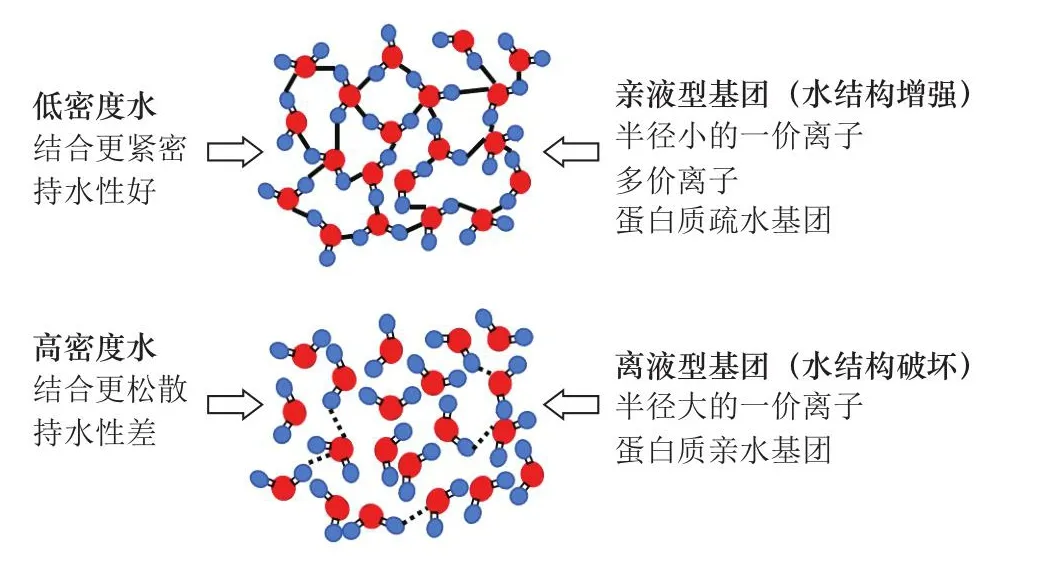
图2 高密度水、低密度水的示意图[7]Fig.2 Illustration of high-density and low-density water[7]
2 肉制品持水性理论
在肉品持水性理论研究过程中,除了蛋白质与其邻近的几层水分子的关系,肌原纤维蛋白网络及基质蛋白网络体系如何维系其中的水分也十分重要。目前肉品持水作用力的基础理论主要包括粗细肌丝之间的静电作用力[3,26]、渗透压[1]和毛细作用力[27]。各个理论侧重点不同,尚无单一理论能完美解释不同条件下的肉品持水现象。
2.1 静电作用力
蛋白质在偏离自身等电点时,表面会携带有净电荷。Hamm[3]指出,肌原纤维蛋白或肌丝之间的静电斥力会造成肌原纤维的溶胀,而肌肉微观结构中一些横向连接(Z线、肌动球蛋白等)会限制肌原纤维的溶胀,与静电斥力达到平衡。在肌纤维层面,肌原纤维与肌纤维膜之间的连接以及肌纤维外部的肌内膜也会限制肌纤维的溶胀。根据Hamm的观点,肉品持水性由静电斥力和横向连接作用共同决定。水分子在肌丝蛋白之间受到吸引(极性基团)和排斥(疏水基团)的共同作用,在肌原纤维网络之中形成一种类似冰结构的水分子簇。静电作用力理论能够很好地解释诸多因素,如pH值、盐含量等对于肉品持水性的影响。
2.2 渗透压
与Hamm的观点略有不同,Offer等[1]认为由于邻近肌丝之间的间距大约在15~20 nm,这一间距太长而不足以产生足够强的静电斥力。他们各提出了基于渗透压的肉品持水理论。Hamm在解释氯化钠对于肉品持水性的促进作用时提出带负电的氯离子会选择性地结合到肌丝上,增加肌丝的负电荷,增大静电斥力;Offer等认同氯离子选择性结合到肌丝,但由于肌丝不能自由移动,静电相互作用会导致带正电荷的离子(即钠离子)向肌丝附近富集,在水相中形成不均匀的离子分布。这一不均匀的离子分布在肌丝之间形成渗透压差异,从而导致水分子进入,引起肌原纤维溶胀。同样地,肌肉结构中的横向桥联限制了肌原纤维的溶胀,与渗透作用达到平衡。
2.3 毛细作用力
肌丝之间的间距很小,这一距离理论上可以产生约300 m水柱的毛细作用力[28],因此,有学者指出毛细作用力是维系肉品持水性的重要因素[4]。但Puolanne等[7]指出肌肉宰后成熟过程中,肌丝间距的减小理论上会导致毛细作用增强,从而增强持水性,然而实际体系持水性却随着肌丝间距的减小而减小。他们指出毛细作用可能仅适用于肌纤维外的水分保持,而且在复杂的肌原纤维网络之中,很难去界定毛细管孔径。
3 持水性的表征方法
持水性的测定通常需要向肉品施加作用力,使水分从体系中释放,这一作用力可以是重力,也可以借助挤压、离心等外力,然后通过测定肌肉中自由水在外力作用下的损失量,表征肌肉的持水性[29]。不同的持水性测定及表征方法基于的原理略有不同,清晰地了解和区分这些不同的持水性测定表征方法对于理解实验中观测到的持水性现象十分重要。
3.1 滴水损失及汁液流失
滴水损失是指肉品在不同环境条件下汁液滴落造成的质量损失。最常见的测定方法是将肉块(约30~100 g)悬挂在袋子中,并在冷藏条件下贮藏1~2 d,确保肉和袋子不接触且方便汁液滴落。最终称量贮藏前后的肉块质量,从而计算得出滴水损失[30]。也可以将肉块切成小块放置在托盘,收集汁液后计算,但肉的表面积会对结果有影响。这一方法因其简便性而被广泛使用。汁液流失与滴水损失类似,指的是肉类在贮藏、零售期间在包装容器中损失的水分。典型的汁液流失量大约占新鲜瘦肉质量的2%~6%,冻融循环、宰前应激(可能导致PSE(pale, soft, exudative)肉)等过程会增大水分流失[1]。
3.2 蒸煮损失
蒸煮损失是常见的肉类持水性指标,对肉块蒸煮前后称质量即可计算蒸煮损失。蒸煮损失受到肉类宰后成熟、蒸煮温度、蒸煮条件等影响,一般不超过40%[1]。在所有表征持水性的方法中,蒸煮损失与肉类的多汁性相关度最高[29]。
3.3 滤纸挤压法
该方法是将肉块切碎,称取固定质量(如300 mg)并放置在滤纸中心,利用重物(如200 g)持续挤压约5 min,计算滤纸前后的质量变化,或者计算滤纸上汁液润湿的面积和肉泥的面积之差,Wierbicki等[31]选取了部分肉类进行研究,得出可被挤出的水分占肉中总水分的30%~50%。该方法很早就已经被使用,近年来有研究人员将图像采集系统和分析系统整合到该方法中,提出了持水性快速测定方法,并命名为WHC trend[32]。
3.4 离心法
离心法是借助离心力分离肉类中的部分水分,需要确保离心损失的水分可以被收集在离心管的底部而不与肉接触,以避免重新被吸收。这一方法简便,并且与汁液损失测定原理类似,同时测定结果相关度高,因此被广泛采用。赫尔辛基大学的Ertbjerg教授研究团队[19]扩展了此方法的应用,发现对提取的肌原纤维进行离心同样可以用来表征不同情形下肉类体系的持水性,每克肌原纤维离心之后截留的水分大约在5~9 g[33-34]。
3.5 肌原纤维溶胀以及肌肉水合能力
3.1~3.4 节中介绍的方法均是通过测定水分损失表征肉品持水性,而也有学者采用肌原纤维的溶胀以及肌肉的水合能力来表征肉品持水性。肌原纤维的溶胀被广泛用作肉品持水性的理论模型。利用相差显微镜可以直接观测提取的肌原纤维的形态,肌原纤维的宽度反映了肌原纤维的横向溶胀状态。而将肉块浸泡在溶液中,称量浸泡前后的质量变化即可反映肉品持水的特性[8]。
3.6 无损检测法
上述持水性测定或表征方法都需要破坏肉类结构,一些快速、无损的方法越来越受到研究人员和肉品行业的欢迎。研究表明低场核磁共振的横向弛豫时间(T2)可以有效地测定肉类中的自由水[13]。Kamruzzaman等[35]采集了猪肉、牛肉、羊肉的高光谱图像,成功建立了预测红肉持水性的模型。Kaewthong等[36]成功地利用了电导率来评价鸡胸肉的持水性。
4 影响肉品持水性的因素
蛋白质净电荷以及肌肉的微观结构是影响持水性的两大重要内在因素[15],肉品生产过程中如宰前应激、胴体冷却、冷藏、冷冻/解冻、腌制等外部因素均可以通过影响蛋白质净电荷或肌肉微观结构而影响其持水性。
4.1 蛋白质净电荷
蛋白质上部分氨基酸残基(Lys、Arg、His、Asp、Glu等)以及碳末端羧基和氮末端的氨基可以发生解离从而携带电荷(图3)。肉类在宰后成熟过程中,随着组织内乳酸的积累,pH值会下降。一般猪肉、牛肉的最终pH值会略高于肌原纤维蛋白整体的等电点(pH 5.0左右)[1,37],因此肌原纤维蛋白整体携带净的负电荷。研究表明,肌丝净电荷的多少影响肌丝间距并最终影响肉品的持水性。图3展示了不同pH值条件对肌丝电荷以及肌原纤维蛋白体系持水性的影响[9]。在pH值接近等电点时,肌丝因含有大致相同量的正负电荷而净电荷接近0。通过加酸加减都可以使pH值偏离等电点而增加肌丝的净电荷(表现为图3中肌原纤维可以结合更多的带电荷染料)。其中pH值增大导致组氨酸的解离平衡主要以不带电荷的形式存在,相反地,pH值减小导致谷氨酸、天冬氨酸等解离平衡时主要以不带电荷的形式存在,两种情形下蛋白质的整体净电荷都会增大,从而增大了肌丝间的静电互斥[26]或者造成反离子的不均匀分布导致渗透压差[1],使得肌丝间距变大,最终增强了持水性。当pH值进一步增大或者降低时,由于体系酸性或碱性过大,导致肌原纤维网络体系崩塌而造成持水性下降。NaCl是肉类加工中常用的添加物,当添加NaCl时,由于氯离子选择性与粗肌丝结合,当肌丝本身携带净负电荷时,氯离子的加入会增加总净负电荷,从而增强静电排斥,肌丝间距增大,持水性升高;相反地,当肌丝携带净正电荷时,氯离子的加入会减少总净电荷,造成持水性下降。由于肌丝净电荷的载体为肌原纤维蛋白,蛋白质理化性质的变化也可能影响净电荷,比如Bao Yulong等[33]在研究模拟氧化体系下肌原纤维的持水性时发现氧化增大了持水性,推测可能是蛋白质组氨酸残基的氧化生成羰基化合物2-oxo-histidine,从而失去携带的正电荷,导致净的负电荷增加。主要肌原纤维蛋白净电荷的增加也在等电点电泳结果中得到证明[33];而肌浆蛋白的变性导致溶解度下降,可能会沉积到肌原纤维蛋白表面从而遮蔽部分电荷[16]。
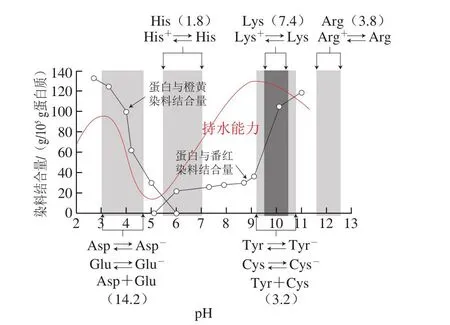
图3 pH值对肌丝电荷以及肌原纤维蛋白体系持水性的影响[9]Fig.3 Effect of pH on myofilament charge and the water-holding capacity of myofibrillar protein[9]
由于pH值和氨基酸基团的pKa值共同决定侧链基团的解离情况,因此pH值是影响肌肉蛋白净电荷从而影响其持水性的重要原因。动物的宰前应激影响肌肉内糖原含量以及糖酵解的速率从而影响最终pH值,如持水性良好DFD肉的形成与宰前糖原过度消耗导致肉类最终pH值偏高有关。而胴体的冷却过慢虽然不改变肌肉最终pH值,却影响pH值下降的速率,如持水性差的PSE肉的形成与胴体pH值下降过快有关[1]。在腌制过程中,一些可食用酸碱[38]、磷酸盐[39]的加入都有可能改变体系的pH值从而改变持水性。在肉品冷冻过程中,也涉及pH值的变化,伴随着肌肉中水分的冻结,溶质浓缩效应导致各类离子(包括H+)浓度增加,从而改变未冻结水分的pH值。Zhang Yuemei等[34]等猜测在冷冻时,H+可能会被部分地束缚在冰晶中从而影响体系局部的pH值,相较于慢速冷冻,快速冷冻可能导致H+更多地被截留在冰晶中,从而未冻结部分pH值更高。此外,冷冻电势可能也是影响冷冻肉类未冻结部分液体pH值变化的原因,冷冻产生的电势会被体系中移动性很强的H+或OH-中和,最终导致局部pH值发生变化[40](图4)。
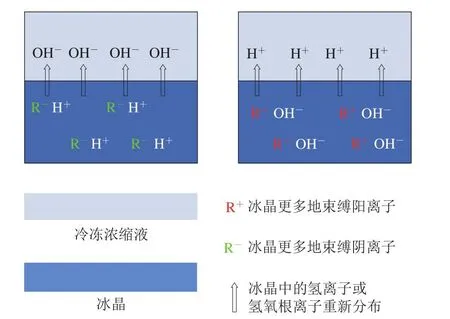
图4 冷冻过程中冰水两相电势介导的pH值变化机理示意图[40]Fig.4 Schematic illustration of pH changes in frozen electrolyte solutions mediated by freezing potential[40]
4.2 微观结构
肌纤维的结构变化是净电荷之外又一影响肌肉持水性的重要因素。肌纤维结构决定了肌肉中水分分布,也限制了肌原纤维/肌纤维的过度溶胀。肌小节是肉类的基本结构单元,宰后肌动球蛋白横桥的形成减少了能容纳的水分,多聚磷酸盐的加入可以破坏肌动球蛋白的连接从而增强持水性[41]。除此之外,肌小节的横向收缩也会导致肌原纤维的体积减小,挤出部分水分[18]。肌原纤维通过肌间线蛋白、纽带蛋白等与细胞膜相连接,在加工或贮藏过程中,如果连接蛋白降解充分,肌原纤维失去与细胞膜的连接,则肌原纤维的收缩将不会联动肌纤维整体收缩,从而减少了水分的流失;如果连接蛋白的降解不充分,则这些连接仍然存在,肌原纤维的收缩可以借助这些连接传递到整个肌纤维,从而使肌细胞直径减小将水分排出细胞外。由于细胞外的水分更容易流失,因此肉品持水性下降(图5)。肉的宰后成熟过程往往伴随着细胞骨架蛋白在内源酶的作用下降解,而连接蛋白的降解减少了收缩的肌原纤维对于肌纤维整体的牵制,肌纤维重新溶胀,肌细胞直径增大使肉品持水性在宰后成熟之后变好[5-6]。Zeng Zhen等[42]研究发现,肌原纤维Z线附近的辅肌动蛋白可以被钙蛋白酶降解,从而导致Z线弱化,减少对肌原纤维横向溶胀的限制,肌原纤维的持水性得以提升(图5)。
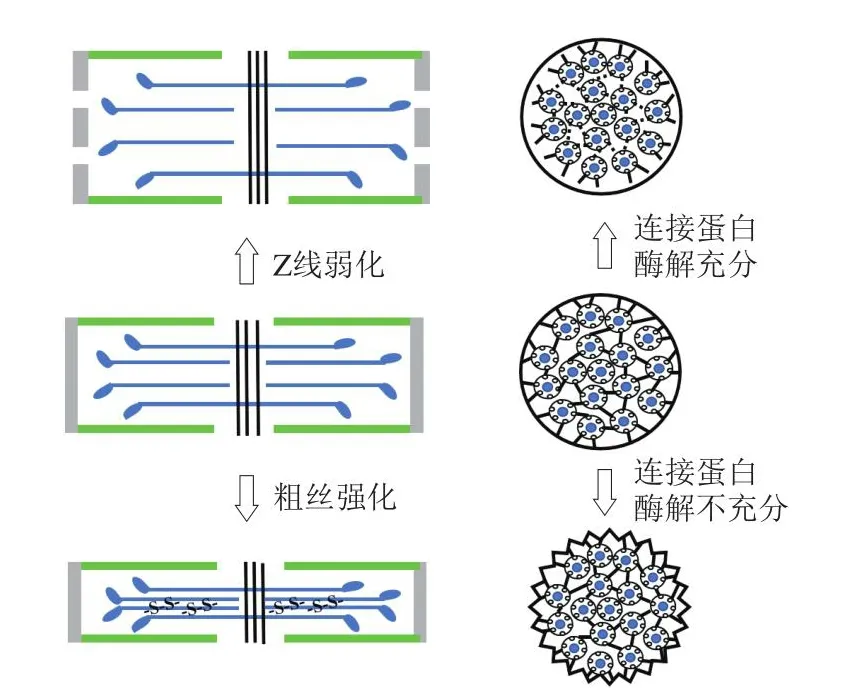
图5 肉品空间位阻效应对持水性影响的机理示意图[6,42-43]Fig.5 Illustration of the mechanism of changes in water-holding capacity of muscle due to steric effect[6,42-43]
生鲜肉在贮藏加工过程中(如含氧包装、辐照、冷冻等),其蛋白质容易发生氧化,而已有的研究表明蛋白的氧化修饰也可以通过影响肉品微观结构从而影响持水性[44]。蛋白质氧化影响肉的持水性主要有两种机制:1)肉品中主要的蛋白水解酶——µ-钙蛋白酶在宰后早期可能因发生氧化而钝化,减弱了其对一些细胞骨架蛋白的降解能力;2)氧化促使蛋白分子间形成二硫键或羰基介导的交联从而强化了粗丝的结构完整性,限制了肌原纤维粗丝的横向溶胀(图5),造成持水性下降[45]。
根据Hamm[26]的观点,肌原纤维蛋白对肉品持水性的贡献约占50%,肌浆蛋白仅占3%左右。最近研究表明肌浆蛋白在特定条件下也参与肉品中微观结构的形成,从而影响持水性[46]。在类PSE肉中,肌浆蛋白的沉积与肌丝间距的减小同时发生,变性的肌浆蛋白在细胞内肌原纤维外以及肌纤维外发生沉积,形成了网络结构,从而部分抵消了肌原纤维蛋白变性对持水性的下降作用[17]。最新的研究也进一步证实了在类PSE肉中,肌浆蛋白的变性提升了肌原纤维的持水性[46]。肌肉中结缔组织的存在也是限制肌肉溶胀的因素,比如完整的肌内膜会限制肌纤维的过度溶胀。目前,肌肉中研究比较成熟的是肌原纤维蛋白以及肌浆蛋白,关于膜蛋白以及其他低丰度的骨架蛋白对于持水性影响的研究比较有限。随着蛋白组学技术在食品科学领域的应用愈加广泛,通过蛋白组学技术可以发现高持水性的生物标志物,从而为研究持水性的调控机制提供更多思路[43,47-51]。Liu Yueyue等[48]研究发现冷冻鳙鱼肉持水性的下降伴随着蛋白质分子的氧化,而最主要的氧化修饰包括与脂肪氧化产物丙二醛和4-羟基壬烯醛(4-hydroxynonenal,HNE)的加成。Zhang Muhan等[50]发现高持水性鹅肉与低持水性鹅肉相比,差异蛋白主要集中在结构蛋白、代谢酶、抗氧化酶、应激蛋白;Zuo Huixin等[51]指出肌球蛋白轻链、热休克蛋白HSP27、磷酸丙糖异构酶可以作为预测牦牛肉高持水性的生物标志物。然而,差异蛋白只能体现含量上的差异,对于蛋白是否变性、酶解程度等指标则很难做出区分,而这些又是影响持水性的重要因素。随着组学技术的进步,上述蛋白变性等问题也将迎来更加全面且合理的解决方案。如已有研究人员开发出限制性酶解-液相色谱-质谱联用分析技术,从而实现对蛋白质结构变化的高通量筛选[52]。
除了上述提及的蛋白酶解、氧化、变性等,其他一些物理性的影响因素也会导致肌肉微观结构发生变化。生鲜肉类的汁液主要从切口处流失,肉块的大小、切口与肌纤维方向的关系、支撑方式(悬挂、或放置在台面)都可能通过改变汁液流失通道的结构从而影响持水性[1]。冷冻/解冻过程冰晶的物理破坏作用也是影响肉品持水性的一大因素。对于慢速冷冻的肉品,冰晶大且主要分布在细胞外,冻藏过程中温度波动造成的重结晶使冰晶体积进一步增加,大冰晶会破坏肉的微观结构,造成解冻时更多的汁液流失[53]。
5 结 语
学术界关于肉品持水性变化机理方面的研究开展较早,各个模型体系下肉品持水的理论基础已经得到了很好地阐明。然而,实际肉品体系中所涉及到的众多理化因素(pH值、离子强度及类型、蛋白酶解、蛋白变性等)大多处于一个动态平衡的过程。肉品体系的一大特点是很难预测任何单一因素对肉品品质产生的确切影响。因此,肉品持水性机理仍然有待进一步阐明。随着蛋白质化学、水的功能特性研究的进步和定量蛋白组学、超分辨显微镜等高新技术的发展,肉制品持水性的理论研究将会取得新的成果。
