分子伴侣共表达对重组蔗糖异构酶异源可溶性表达的影响
2022-11-30孙佳明高向红李明玉王从纲李宪臻
孙佳明, 庞 焦, 高向红, 李明玉, 王从纲, 李宪臻
(大连工业大学 生物工程学院,辽宁 大连 116034)
蔗糖异构酶主要来源于细菌,已报道的多种产蔗糖异构酶的细菌包括大黄欧文菌(Erwinia rhapontici)、沙雷氏杆菌(Serratia plymuthica)、克雷伯式杆菌 (Klebsiella sp.)和多源发散菌(Pantoea dispersa)等[9]。野生菌株普遍产酶水平低,难以达到工业化生产应用的要求,如Erwinia rhapontici NX-5发酵酶活力为1.3 U/mL[10],优化后的Klebsiella sp.LX3的发酵酶活力达到15.12 U/mL[11]。利用原核系统对重组蛋白质进行异源表达是提高其产量的有效手段,具有生产成本低、工艺操作简便、生产效率高等多种优势。目前针对蔗糖异构酶的异源表达,采用大肠杆菌为表达宿主的研究工作最多,如源自沙雷氏杆菌 (Serratia plymuthica)、多源发散菌(Pantoea dispersa UQ68J) 和 克 雷 伯 式 杆 菌(Klebsiella sp.UQ14S)的蔗糖异构酶在大肠杆菌中实现了胞内表达[12-13],然而,蔗糖异构酶的可溶性表达水平很低,主要以包涵体形式表达,严重制约了蔗糖异构酶的基础研究和应用[14-15]。针对这一问题,有研究者采用融合谷胱甘肽巯基转移酶(glutathione S-transferase,GST)标签或高密度发酵手段在一定程度上提高了蔗糖异构酶的可溶性表达量,但改善程度仍然有限[16-18]。此外,通过优化诱导表达条件,如调整诱导剂的加入量、改变诱导温度在很多情况下对目的蛋白质在大肠杆菌中可溶性表达的提高有限,而关于蛋白质折叠的研究为提高重组蛋白质的可溶性表达提供了新思路,研究表明不同分子伴侣可以在目的蛋白质多肽链翻译后的各个阶段发挥作用,通过不同机制促进目的蛋白质正确折叠从而提高其可溶性表达水平,如通过共表达分子伴侣质粒pGro7使核糖核酸外切酶R的可溶性表达量提高了54.50%[19]。通过比较4种分子伴侣质粒pGKJE8、pGro7、pG-Tf2、pTf16对牛支原体膜蛋白M1的截断片段在大肠杆菌中表达的影响,发现分子伴侣质粒pG-KJE8和pG-Tf2能显著提高M1截短片段的可溶性表达[20]。目前,关于蔗糖异构酶异源高效可溶性表达的研究工作较少,尤其是缺少利用分子伴侣共表达策略调控其可溶性表达的研究工作。
在作者前期工作中,已分离到了Klebsiella sp.LX3菌株,该菌株生产的蔗糖异构酶具有很高的蔗糖转化率(99.5%),其转化蔗糖的主产物为异麦芽酮糖,具有潜在的工业应用价值[11,16]。尽管前期工作中对该菌株表达的蔗糖异构酶通过融合GST标签在一定程度上改善了SIase的可溶性表达量,但改善程度有限,并且后续纯化需要蛋白酶切除标签和凝胶过滤层析得到纯酶[16]。同时,其他文献中也报道了多种不同来源的蔗糖异构酶可溶性表达水平较低,主要以包涵体形式表达[14-15]。因此,为了提高蔗糖异构酶的异源表达水平,作者以源自Klebsiella sp.LX3的SIase为研究对象,构建重组表达载体pET-24b-SIase,转入大肠杆菌E.coli BL21(DE3)中进行表达,进一步将4种分子伴侣质粒(pKJE7、pGro7、pG-Tf2、pTf16)分别与基因SIase在大肠杆菌E.coli BL21(DE3)中共表达,通过辅助蛋白质正确折叠减少包涵体产生并促进SIase可溶性表达。进一步将筛选所得含有最佳分子伴侣的工程菌进行培养,利用金属离子螯合层析技术纯化可溶性SIase,并进行酶学性质表征和产物特异性分析,从而为规模化制备SIase的应用基础研究提供参考,为SIase在工业上的大规模应用奠定基础。
1 材料与方法
1.1 材料与试剂
菌株E.coli BL21(DE3)、E.coli DH10B、质粒pET-28a-SIase:作者所在实验室保藏;卡那霉素、氯霉素、L-阿拉伯糖、四环素、异丙基-β-D-硫代半乳糖苷(isopropyl-beta-D-thiogalactopyranoside,IPTG)、SanPrep柱式质粒DNA小量抽提试剂盒、SanPrep柱式DNA胶回收试剂盒、Ni-NTA 6FF琼脂糖纯化树脂、咪唑、BCA蛋白质定量检测试剂盒:生工生物工程(上海)股份有限公司产品;分子伴侣质粒(pKJE7、pGro7、pG-Tf2、pTf16)、DNA和蛋白质相对分子质量标准品、PrimeSTAR®HS DNA Polymerase:宝生物工程(大连)有限公司产品;异麦芽酮糖:Sigma公司产品;其他化学试剂:天津市科密欧化学试剂有限公司产品。
LB液体培养基:10 g/L胰蛋白胨、5 g/L酵母粉、10 g/L NaCl,pH 7.2;LB固体培养基为在LB液体培养基中加入15 g/L琼脂;LLB培养基:10 g/L胰蛋白胨、5 g/L酵母粉、5 g/L NaCl,pH 7.2。Buffer A缓 冲 液:0.5 mol/L氯 化 钠、50 mmol/L磷 酸 二 氢钠-氢氧化钠,pH 8.0;Buffer B缓冲液:0.5 mol/L氯化钠、50 mmol/L磷酸二氢钠-氢氧化钠、0.03 mol/L咪唑,pH 8.0;Buffer C缓冲液:0.5 mol/L氯化钠、50 mmol/L磷酸二氢钠-氢氧化钠、0.05 mol/L咪唑,pH 8.0;Buffer D缓 冲 液:0.5 mol/L氯 化 钠、50 mmol/L磷酸二氢钠-氢氧化钠、0.10 mol/L咪唑,pH 8.0;Buffer E缓冲液:0.5 mol/L氯化钠、50 mmol/L磷酸二氢钠-氢氧化钠、0.20 mol/L咪唑,pH 8.0。
1.2 仪器与设备
Milli-Q超纯水过滤系统:美国密理博Millipore公司产品;HITACHI-CR21GⅢ超速冷冻离心机:日立(中国)有限公司产品;TGL-16高速台式冷冻离心机:湘仪离心机仪器有限公司产品;通用型垂直电泳仪:上海生工生物科技有限公司产品;Infinite F50酶标仪:瑞士帝肯Tecan公司产品;PCR仪、电转仪:德国Eppendorf公司产品;NanoVue Plus超微量紫外分光光度计、ImageQuant LAS4000凝胶成像分析系统:美国通用电气(中国)有限公司产品;高效液相色谱分析仪 (Agilent Technologies 1260 Infinity):美国安捷伦科技公司产品;超声波细胞粉碎机:宁波新芝生物科技股份有限公司产品;紫外分光光度计:上海元析仪器有限公司产品。
1.3 方法
1.3.1 SIase表达工程菌的构建为了便于重组SIase的分离纯化,并且尽可能减少在SIase中引入额外的氨基酸残基,作者以所在课题组前期构建的携带源自Klebsiella sp.LX3的SIase基因的质粒pET-28a-SIase为模板(SIase基因存在于BamH I和Xho I酶切位点之间),通过PCR扩增和酶切连接技术将SIase基因扩增并插入到pET-24b载体的Nde I和Xho I酶切位点之间,从而在SIase的C端引入组氨酸标签,并且N端不会引入多余的氨基酸残基。首先设计引物进行PCR扩增含酶切位点的SIase基 因 片 段。上 游 引 物F:5′-GAGTGATC CATATGGCACCATCCTTGAATCAGGATATTCACG-3′,下游引物R:5′-CTACTGCTCGAGCCGCAGCTTA TACACACCTGCC-3′,其中下划线标识分别为限制性核酸内切酶QuickCutTMNde I和QuickCutTMXho I的识别位点。PCR反应体系为:5×Prime STAR®HSBuffer 10 μL,DNTP 4 μL,上游引物2 μL,下游引物2 μL, 模 板 质 粒 加 入2 μL,PrimeSTAR®HS Polymerase 0.5 μL,灭菌双蒸水补至50 μL。PCR扩增产物用琼脂糖凝胶电泳和胶回收试剂盒进行回收。将回收得到的PCR产物和pET-24b载体用QuickCutTMNde I和QuickCutTMXho I进行双酶切。酶切反应体系为PCR胶回收产物1 000 ng,10×Green Buffer 2 μL,QuickCutTMNde I和QuickCutTMXho I各1 μL,灭菌双蒸水补至20 μL,质粒载体1 000 ng,10×Green Buffer 2 μL,QuickCutTMNde I和QuickCutTMXho I各1 μL,灭菌双蒸水补至20 μL;酶切反应条件为37℃反应30 min。酶切产物经琼脂糖凝胶电泳分离后利用DNA胶回收试剂盒回收线性化载体质粒和目的基因片段,继续用连接酶进行连接反应。将连接产物热转化至E.coli DH10B感受态细胞,涂布于含终质量浓度为50 μg/mL卡那霉素的LB固体平板上。挑取单菌落进行菌落PCR鉴定,筛选到的阳性单克隆送吉林省库美生物科技有限公司进行测序,测序正确的重组质粒命名为pET-24b-SIase,将其转入E.coli BL21(DE3)感受态细胞中得到工程菌。
1.3.2 分子伴侣与SIase共表达工程菌的构建分别 将4种 分 子 伴 侣 质 粒pGro7、pKJE7、pG-Tf2、pTf16(见表1)转入E.coli BL21(DE3)感受态细胞中,利用含有终质量浓度为30 μg/mL氯霉素的平板筛选阳性克隆,并且再次将含有分子伴侣的重组菌制备成感受态细胞,转入pET-24b-SIase质粒,涂布于含有氯霉素和卡那霉素的平板进行筛选,得到同时含有分子伴侣质粒和pET-24b-SIase质粒的工程菌。
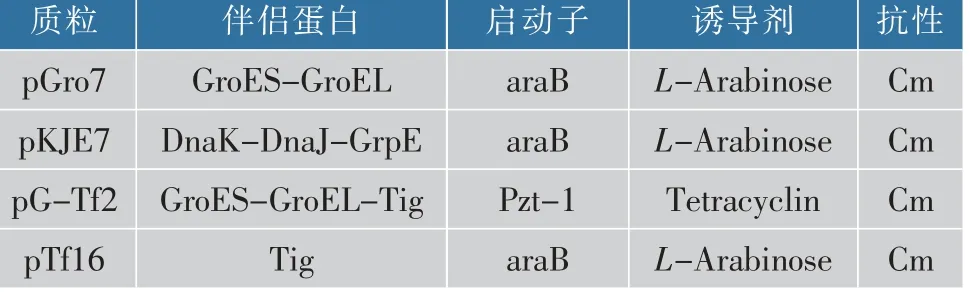
表1 携带分子伴侣质粒信息Table 1 Information of plasmids harboring different molecular chaperons
1.3.3 SIase的诱导表达挑取含有SIase基因的阳性单克隆接种于5 mL含终质量浓度为50 μg/mL卡那霉素的LB液体培养基中,37℃、200 r/min培养14 h得到种子液。将种子液以1%(体积分数)比例转入含50 μg/mL卡那霉素的LLB液体培养基中进行扩大培养,37℃、200 r/min培养至菌体OD600达 到0.6~0.8时,加 入 终 浓 度 为0.5 mmol/L的IPTG,在16℃、200 r/min条件下培养20 h进行诱导表达。共表达工程菌需要进行分子伴侣与SIase的分别诱导,首先挑取共表达工程菌的阳性单克隆接种于5 mL含50 μg/mL卡那霉素和30 μg/mL氯霉素的LB液体培养基中,于37℃、200 r/min培养14 h得到种子液。将种子液以1%(体积分数)比例转入含50 μg/mL卡那霉素和30 μg/mL氯霉素的LLB液体培养基中,同时加入诱导剂10 μg/mL的四环素(pG-Tf2)或500 μg/mL L-阿拉伯糖(pGro7、pKJE7、pTf16)以诱导分子伴侣表达,继续在37℃、200 r/min培养至菌体OD600达到0.6~0.8时,加入终浓度为0.5 mmol/L的IPTG,在16℃、200 r/min条件下培养20 h进行诱导表达。将未进行和进行了共表达的工程菌分别于8 000 r/min、5 min条件下离心收集菌体细胞,加入Buffer A缓冲液重悬,在冰水浴中超声破碎(功率500 W,破碎5 s,间隔3 s,破碎30 min),10 000 r/min离心15 min分离可溶性上清液和沉淀组分,之后取20 μL可溶性上清液与5 μL 5×上样缓冲液混合,将破碎后沉淀用Buffer A缓冲液重悬后取20 μL与5 μL 5×上样缓冲液混合,上述样品经沸水处理5 min后分别吸取10 μL上清液样品和5 μL沉淀样品加至胶孔中进行SDS-PAGE检测。
为此,工程项目划分必须坚持由项目法人组织监理、设计及施工等单位进行,并确定主要单位工程、主要分部工程、重要隐蔽单元工程和关键部位单元工程。项目法人在主体工程开工前将项目划分表及说明书面报相应质量监督机构确认。工程实施过程中,需对单位工程、主要分部工程、重要隐蔽单元工程和关键部位单元工程的项目划分进行调整时,项目法人要重新报送工程质量监督机构进行确认。
1.3.4 重组SIase的分离纯化将筛选到含最佳分子伴侣质粒的工程菌进行诱导表达,于8 000 r/min、5 min条件下离心收集菌体细胞,加入Buffer A缓冲液重悬,在冰水浴中超声破碎(功率500 W,破碎5 s,间隔3 s,破碎30 min),10 000 r/min离心15 min分离可溶性上清液和沉淀组分。由于重组SIase的C端带有组氨酸标签,因此利用金属离子螯合层析法进行纯化。将可溶性上清液经过0.22 μm滤膜过滤后与提前用Buffer A缓冲液平衡的Ni-NTA 6FF琼脂糖纯化树脂结合,流量为1 mL/min。之后分别用10倍柱体积的Buffer A缓冲液、Buffer B和Buffer C缓冲液进行洗杂。最后利用Buffer D和Buffer E洗脱目的蛋白质。将纯化得到的不同组分取20 μL与5 μL 5×上样缓冲液混合,经沸水处理5 min后分别吸取10 μL进行SDS-PAGE检测,纯化得到的目的蛋白质SIase经透析处理后进行蛋白质浓度测定和酶学性质表征。蛋白质浓度采用BCA蛋白质定量检测试剂盒测定。
1.3.5 SIase酶活力的测定100 μL SIase(0.08 mg/mL)的酶液与400 μL含40 g/L蔗糖的磷酸氢二钠-柠檬酸缓冲液(50 mmol/L,pH 6.0)混匀,于特定温度下水浴反应15 min,之后将样品经100℃沸水处理15 min终止反应。将含有反应产物的混合物于10 000 r/min离心15 min,取上清液经0.22 μm滤膜过滤后利用HPLC进行检测。色谱柱为Hypersil APS-2氨基柱,流动相为含有80%(体积分数)乙腈的水溶液,流量为1.0 mL/min。酶活力单位的定义:以蔗糖为底物,每分钟释放1 μmoL异麦芽酮糖所需的酶量为1个酶活力单位(U)。
1.3.6 温度对SIase酶活力的影响取100 μL含0.08 mg/mL SIase的酶液与400 μL含40 g/L蔗糖的磷酸氢二钠-柠檬酸缓冲液(50 mmol/L,pH 6.0)混匀。分别在20、25、30、35、40、45、50、55、60℃水浴反应15 min,之后经灭酶处理后利用HPLC检测反应产物中的异麦芽酮糖并进一步计算酶活力。相对酶活力定义为以测定的最高酶活力为100%计算所得比值为相对酶活力。
1.3.7 pH对SIase酶活力的影响取100 μL含0.08 mg/mL SIase的酶液分别与400 μL含40 g/L蔗 糖 的pH为4.0、4.5、5.0、5.5、6.0、6.5、7.0、7.5、8.0的磷酸氢二钠-柠檬酸缓冲液混匀。在最适温度条件下水浴反应15 min,之后经灭酶处理后利用HPLC检测反应产物中的异麦芽酮糖并进一步计算酶活力。相对酶活力定义为以测定的最高酶活力为100%计算所得比值为相对酶活力。
1.3.8 SIase的动力学参数测定分别以不同浓度的 蔗 糖 溶 液(8、16、32、40、80、160、200、320、400、640 mmol/L)为底物进行蔗糖异构酶活力的测定。反应条件为:最适温度和最适pH反应条件下,水浴反应15 min。反应结束后将样品经100℃沸水灭酶15 min,将离心得到的上清液经0.22 μm滤膜处理后进行HPLC检测分析。利用GraphPad Prism5.0基于Michaelis-Menten方程进行非线性拟合得到动力学参数Km、Vmax,计算得到kcat、kcat/Km。
1.3.9 SIase的产物特异性分析取100 μL含0.08 mg/mL SIase的酶液与400 μL含40 g/L蔗糖的磷酸氢二钠-柠檬酸缓冲液(50 mmol/L,pH 6.0)混匀,分别在30、35、40℃下水浴12 h进行转化反应,之后将反应样品经100℃沸水灭酶处理,10 000 r/min离心15 min得到的上清液经0.22 μm滤膜过滤,之后进行HPLC检测。
2 结果与分析
2.1 SIase在E.coli BL21(DE3)中的重组表达
首先以作者所在实验室保存携带SIase基因的pET-28a表达载体为模板,利用PCR技术扩增得到含酶切位点的SIase基因,与pET-24b载体质粒用QμickCutTMNde I和QμickCutTMXho I双酶切分别获得有黏性末端的SIase基因片段和线性化载体质粒,之后用T4 DNA连接酶进行连接反应并转化进入E.coli DH10B感受态细胞,利用终质量浓度为50 μg/mL卡那霉素的LB固体平板筛选得到单菌落用于菌落PCR鉴定。利用pET系列载体的T7通用引物进行菌落PCR鉴定,如图1所示,结果显示PCR产物在1 500~2 250 bp处存在单一条带,与pET-24b-SIase预期扩增条带1 896 bp位置一致,表明其为阳性克隆。将验证正确的菌液送吉林省库美生物科技有限公司进行基因测序和进一步比对,表明pET-24b-SIase表达载体构建成功。
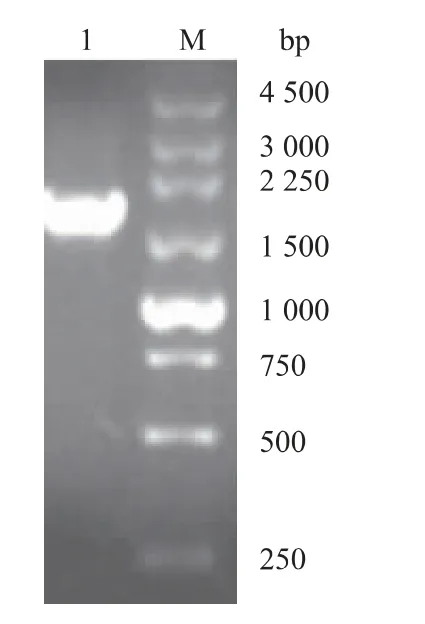
图1 转化重组质粒工程菌的菌落PCR鉴定Fig.1 PCR amplification for identification of gene engineering bacteria transformed with recombinant plasmid
将重组质粒pET-24b-SIase转化至E.coli BL21(DE3)中获得工程菌,对重组工程菌在16℃、诱导剂IPTG终浓度为0.5 mmol/L条件下诱导表达20 h。收集菌体经超声破碎后通过离心分离获得可溶性目的蛋白质上清液和不溶性沉淀,利用SDSPAGE进行检测。如图2所示,经诱导后在约66 400处出现与SIase理论相对分子质量68 140相符的目 的 条 带, 这 与 源 自P.dispersa UQ68J、K.pneumoniae NK33-98-8、E.rhapontici NX-5、S.plymuthica AS9的蔗糖异构酶在大肠杆菌中异源表达的相对分子质量相近[10,12-13,21]。并且SIase主要以包涵体形式存在于不溶性沉淀组分中,表明利用原核表达系统对SIase进行异源可溶性表达存在一定困难。
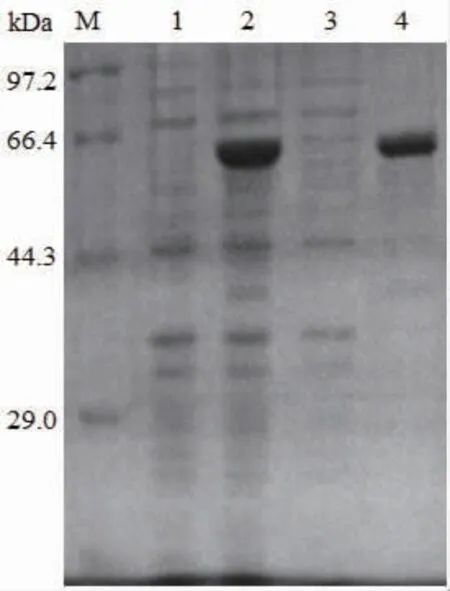
图2 重组SIase的SDS-PAGE分析Fig.2 Analysis of recombinant proteins of SIase by SDSPAGE
2.2 不同分子伴侣对SIase可溶性表达的影响
分子伴侣能分别或通过协同作用参与蛋白质的折叠过程,有利于促进蛋白质正确折叠和可溶性表达[22-23]。为提高SIase的异源可溶性表达水平,通过引入不同分子伴侣质粒实现不同分子伴侣与SIase的共表达,以探究利用分子伴侣共表达策略提高SIase可溶性表达的可行性。将重组质粒pET-24b-SIase分别转化至含有4种不同分子伴侣质粒的E.coli BL21(DE3)感受态细胞中,构建出含有分子伴侣质粒(pGro7、pKJE7、pG-Tf2、pTf16)的共表达工程菌E.coli BL21(DE3)/pET-24b-SIase,进一步发酵培养并利用诱导剂L-阿拉伯糖或四环素和IPTG诱导表达,收集菌体经超声破碎后获得上清液和不溶性沉淀,利用SDS-PAGE进行检测。结果如图3所示,含有分子伴侣质粒pGro7和pG-Tf2的菌体破碎上清液中,可溶性SIase的含量明显提高,其中共表达分子伴侣质粒pGro7改善效果最好。因此,结果表明利用共表达分子伴侣策略能促进SIase的可溶性表达,并且筛选到最佳分子伴侣质粒pGro7,摇瓶发酵酶活力为14.8 U/mL,相比未进行共表达的工程菌发酵酶活力3.5 U/mL有明显提高,并且与已报道Erwinia rhapontici NX-5菌株发酵酶活力1.3 U/mL和Erwinia sp.D12经培养基优化的发酵酶活力10.84 U/mL有一定程度的提高,与优化发酵后的Klebsiella sp.LX3的发酵酶活力为15.12 U/mL相近,从而为进一步系统优化发酵条件打下了基础[10-11,22]。目前大肠杆菌中主要存在3种分子伴侣组,其中DnaK-DnaJ-GrpE和TF因子在新生肽链的早期折叠阶段保护其免于错误折叠和聚集,GroES-GroEL主要协助部分折叠的多肽链克服折叠动力学屏障[23-24]。同时,pG-Tf2分子伴侣组除表达GroES、GroEL外还表达TF因子,但相比pGro7并未产生更好的效果。因此,结果表明影响SIase正确折叠和可溶性表达的关键在于部分折叠的多肽链跨越动力学屏障存在困难,这为后续进一步提高SIase的异源可溶性表达提供了研究思路。
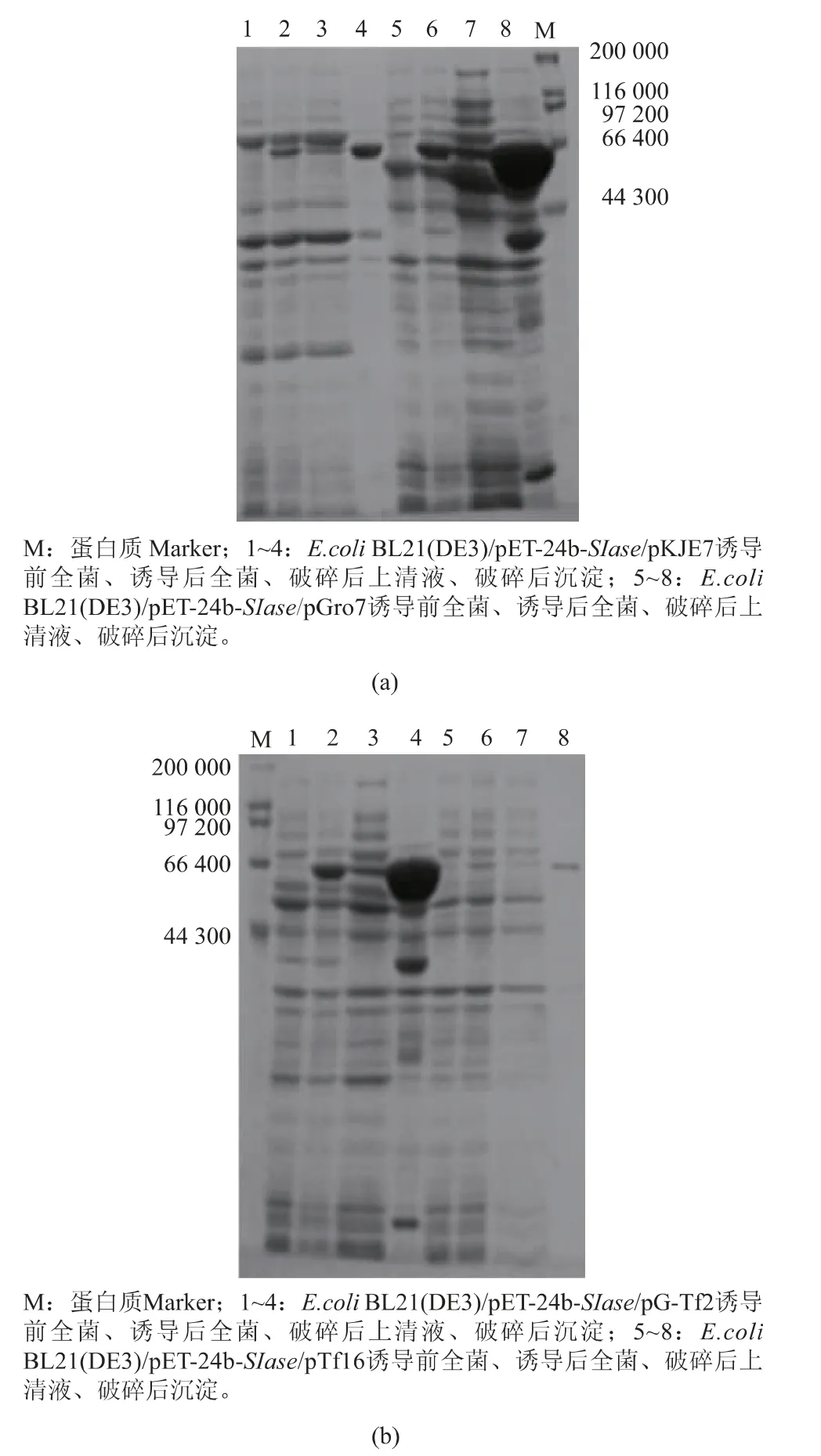
图3 分子伴侣与SIase共表达工程菌的SDS-PAGE分析Fig.3 SDS-PAGE analysis of engineered bacteria coexpressing molecular chaperones and SIase
2.3 重组SIase的分离纯化和酶学性质
由于重组SIase的C端带有组氨酸标签,因此利用Ni-NTA 6FF琼脂糖纯化树脂对含有分子伴侣质粒pGro7的E.coli BL21(DE3)/pET-24b-SIase工程菌细胞破碎上清液中的重组SIase进行纯化,结果如图4所示,利用含有30、50 mmol/L的咪唑的缓冲液有效除去了杂蛋白质,并且利用含100 mmol/L咪唑的缓冲液成功纯化到SIase。本研究中采用分子伴侣共表达策略有效提高了SIase的可溶性表达水平,从而实现了用金属离子螯合层析法一步纯化到纯酶,而已报道研究中采用融合GST标签改善SIase的可溶性表达,后续纯化需要蛋白酶切除标签和凝胶过滤层析才能得到纯酶[16]。因此,本研究中在提高SIase可溶性表达的同时简化了其分离纯化步骤,为针对SIase的基础研究提供了更加简便的方法。
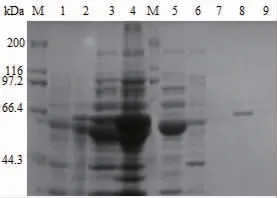
图4 SIase纯化的SDS-PAGE分析结果Fig.4 SDS-PAGE analysis results of SIase purification
将重组SIase纯酶透析后进行酶学性质表征。通过检测SIase在20~60℃的酶活力以确定最适温度。结果如图5所示,SIase在20~40℃下酶活力随温度的升高而增加,在40℃表现出最高酶活力,其中在30~50℃下具有超过70%的相对酶活力,高于50℃后酶活力迅速下降,这一性质与已报道重组蔗糖异构酶的最适温度主要在30~50℃相似[10,12]。
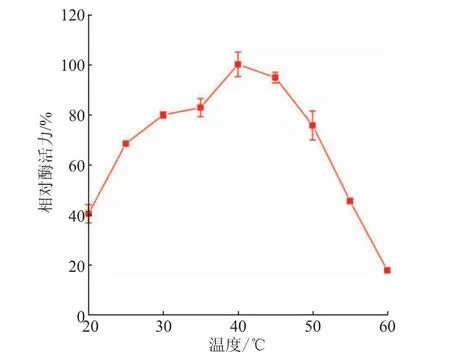
图5 温度对SIase酶活力的影响Fig.5 Effect of temperature on enzymatic activity of SIase
测定了在pH 4.0~8.0的酶活力,结果如图6所示,在pH 5.0~6.5时SIase具有80%以上的相对酶活力,其中在pH 6.0时具有最高酶活力,而pH低于5.0和高于6.5时,酶活力迅速降低。在最适反应条件下测定SIase的比活力为(317.6±15.6)U/mg。
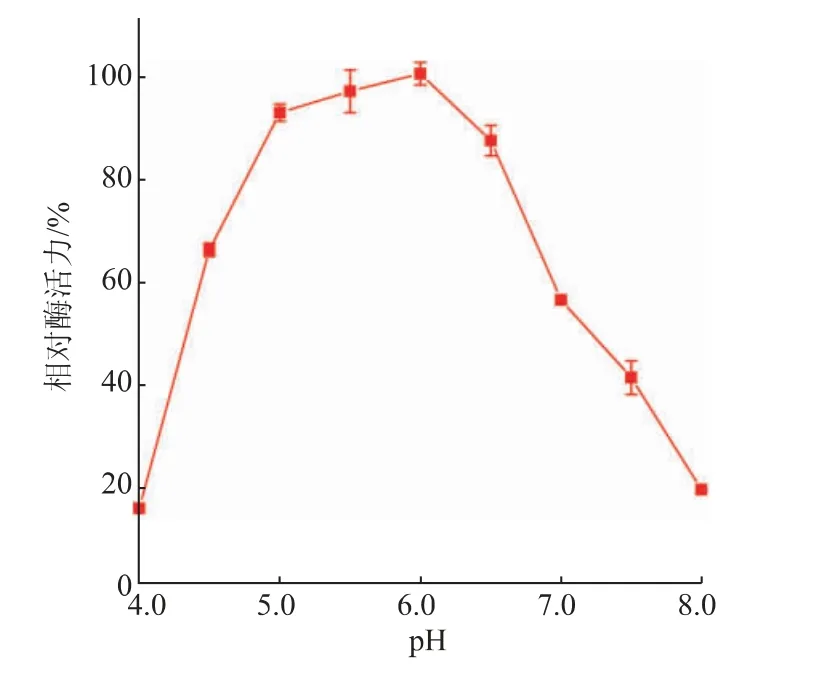
图6 pH对SIase酶活力的影响Fig.6 Effect of pH on enzymatic activity of SIase
在最适反应条件下测定重组SIase对不同浓度底物蔗糖的催化反应初速度,基于Michaelis-Menten方程进行非线性拟合得到动力学参数。结果如表2所示,重组SIase对底物蔗糖的Km为(179.10±20.65)mmol/L、Vmax为 (13 497.0±620.7)μmol/(L·min)、kcat为 (974.4±129.0)s-1,kcat/Km为(5.44±0.72)L/(mmol·s)。

表2 SIase的动力学参数Table 2 Kinetic parameters of SIase
2.4 重组SIase的产物特异性分析
采用HPLC检测了重组SIase的蔗糖转化率为98%,与文献报道不同来源的蔗糖异构酶的蔗糖转化率(99.0%~99.7%)相近[10-11,25],对30、35、40℃下SIase转化蔗糖的反应产物中不同糖组分的含量进行检测,并计算产物中各组分相对含量,结果如表3所示。随着温度提高,重组SIase转化产物中异麦芽酮糖和海藻酮糖比例下降,单糖比例提高,与已报道的重组SIase表现出类似的随温度提高的产物组成变化[16,21]。
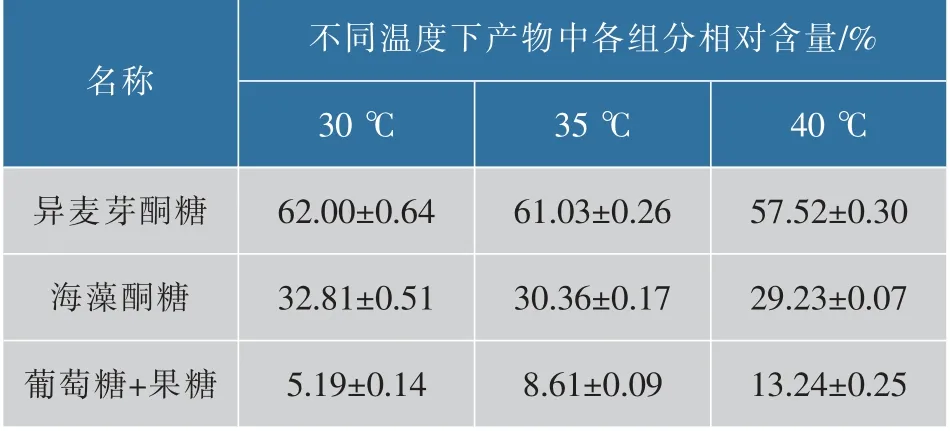
表3 不同温度下SIase催化糖产物组分的分析Table 3 Analysis of the catalytic products compositions of sucrose isomerase at different temperatures
3 结语
作者将源自Klebsiella sp.LX3的SIase在大肠杆菌中进行异源表达,针对其主要以包涵体表达的问题,通过比较4种不同分子伴侣共表达系统对重组SIase可溶性表达的影响,筛选到最佳分子伴侣质粒pGro7,并以此为基础利用金属离子螯合层析技术纯化到了SIase纯酶。重组SIase转化蔗糖的主产物为异麦芽酮糖,其最适温度为40℃、最适pH为6.0。动力学参数Km为(179.10±20.65)mmol/L、Vmax为 (13 497.0±620.7)μmol/(L·min)、kcat为(974.4±129.0)s-1、kcat/Km为(5.44±0.72)L/(mmol·s)。综上,作者利用分子伴侣共表达策略提高了SIase的异源可溶性表达水平,为针对其高效表达的进一步研究工作提供了必要的理论基础和实验依据。但需要注意的是,蔗糖异构酶作为一种食品酶制剂,在大肠杆菌中表达存在一定安全隐患,因此后续工作可以参考本研究中采用的分子伴侣共表达策略,利用食品安全级微生物构建高效异源表达系统,促进蔗糖异构酶在食品工业中的规模化制备和应用。
