重庆地区结核分枝杆菌异烟肼和丙硫异烟胺耐药及其交叉耐药相关基因突变研究
2022-06-08朱大冕刘文果余锋平孔珏颖
朱大冕,刘文果,沈 静,余锋平,孔珏颖,冯 鑫,胡 彦
耐多药结核病(MDR-TB)是指体外药敏试验证实对利福平和异烟肼同时耐药的结核病。根据2020年世界卫生组织报告,2019年全球约有39万例MDR-TB患者,我国约有5.46万例[1]。异烟肼(isoniazid,INH)是结核病标准化疗方案的核心药物[2]。结核分枝杆菌(MTB)对INH耐药后,多采用丙硫异烟胺(protionamide,Pto)代替INH进行治疗。乙硫异烟胺(ethionamaide,Eto)/Pto用于MDR-TB治疗,是目前WHO推荐的MDR-TB治疗药物之一[3]。异烟肼和Eto/Pto的作用机制是抑制MTB分枝酸的合成[2],两药存在部分交叉耐药机制[4],inhA基因及其调控区域突变就是其主要分子机制之一[5]。
本研究将探讨MTB对INH与Pto的耐药情况,采用序列分析的方法检测INH耐药相关基因,分析INH与Pto交叉耐药相关基因inhA突变特点,为耐药基因型检测奠定基础,也为临床治疗MDR-TB提供参考依据。
1 材料与方法
1.1 一般资料 选用2014年9月至2019年3月期间来源于重庆39个区县的233株INH耐药MTB临床分离株以及5株全敏感MTB;中国疾病预防控中心结核病预防控制中心国家结核病参比实验室提供标准株H37Rv。
1.2 试剂 分离培养、菌种鉴定、药敏试验所用试剂均购于珠海贝索生物技术有限公司。培养基内药物终浓度分别为:INH 0.2 μg/mL,RFP 40 μg/mL,SM 4 μg/mL,EMB 2 μg/mL,Ofx 4 μg/mL,Km 30 μg/mL,Cm 40 μg/mL,Pto 40 μg/mL,PAS 1 μg/mL,AK 30 μg/mL。
1.3 药敏试验及鉴定 药敏试验采用比例法,将稀释好的菌悬液分别接种于对照、含药及PNB和TCH培养基,37 ℃培养3~4周。根据耐药百分比判读:耐药百分比=(含药培养基生长的菌落数/对照培养基生长的菌落数)×100%。若耐药百分比大于1%,则认为受试菌对该抗结核药耐药。
1.4 DNA提取 取适量新鲜菌苔于0.5 mL去离子水,85 ℃金属浴30 min,12 000 r/min离心5 min,弃上清;0.5 mL TE缓冲液,100 ℃金属浴20 min;12 000 r/min离心5 min,取上清即为DNA模板。
1.5 基因扩增 耐药相关基因测序使用引物见表1。引物由北京擎科生物技术有限公司合成。反应体积为50 μL,其中DNA 4 μL,Master MIX 25 μL,引物各2 μL,其余由17 μL灭菌蒸馏水补足。扩增条件为:98 ℃预变性2 min;98 ℃ 30 s,退火 30 s(温度根据引物各不同)(表1),72 ℃ 40 s,35个循环;72 ℃延伸5 min。

表1 耐药相关基因测序使用引物Tab.1 Primers used for sequencing drug resistance associated genes
1.6 测序及结果分析 采用1.5%琼脂糖凝胶电泳鉴定PCR产物,送北京擎科生物技术有限公司双向测序。测序结果与NCBI上的katG、inhA、ahpC、kasA基因标准序列进行比对,运用MEGA 6.0进行突变分析。
1.7 统计学分析 采用SPSS 20.0软件分析数据。计数资料比较用χ2检验,P<0.05为差异有统计学意义。
2 结 果
2.1 药敏结果 共收集到233株IHN耐药的结核分枝杆菌临床分离株,其中37株与Pto交叉耐药,交叉耐药率(15.8%)。233株菌株中MDR-MTB有194株(83.2%), 前广泛耐药结核分枝杆菌(pre-XDR-MTB:在对INH和RFP耐药的基础上,同时对二线注射类药物或氟喹诺酮类药物中的一种药物耐药)有82株(35.2%),广泛耐药结核分枝杆菌(XDR-MTB)有8株(3.4%)。菌株来源患者中,男性173例(74.2%)、女性60例(25.8%),患者平均年龄为46.3岁。详见表2。
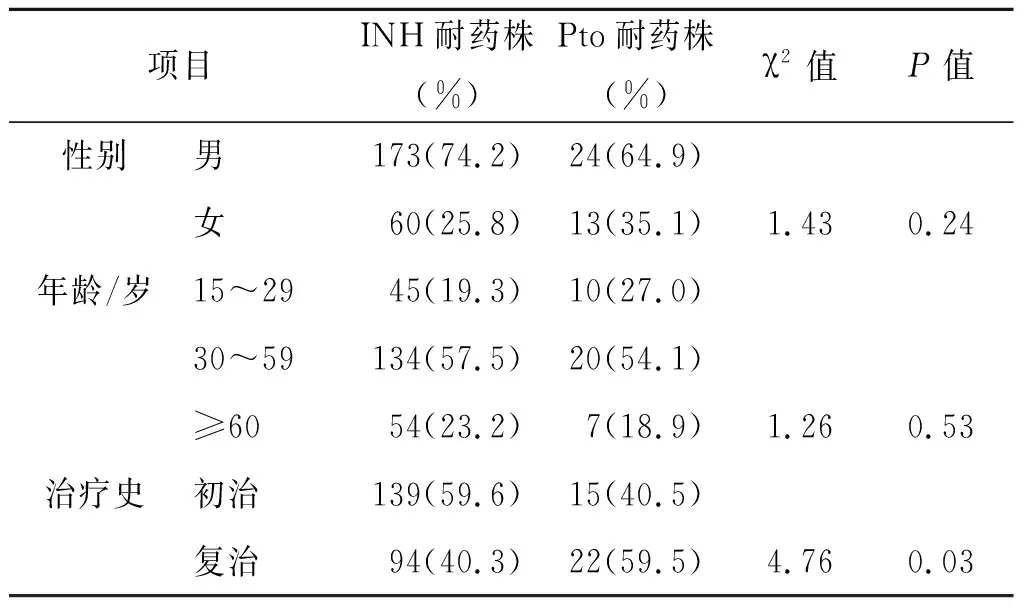
表2 各因素对结核分枝杆菌INH和 Pto耐药的影响Tab.2 Effects of various factors on the resistance of MTB to INH and Pto
2.2 耐药基因测序结果katG突变情况:标准株及5株全敏感菌株均未检测到katG突变。233株耐异烟肼菌株中亦未有katG完全缺失。223株 (96.5%)耐药株katG存在点突变、插入,其中 2个突变位点未见报道(D448G,D419H)。其中195株 (83.6%) 315位点突变 (S315T 188株,S315N 4株,S315I 1株,S315R 2株);189株 (81.1%)463位点突变(R463L),166株 (71.2%)与315位点联合突变。其他位点突变4株,包括D448G 1株、D419H 1株、2株同义突变。另1株在位点1180/1181插入CG。
inhA突变结果:标准株及5株全敏感菌株均未检测到inhA突变。233株INH耐药株中11株(4.7%)inhA发生点突变,包括S94A 1株、I194T 1株、C-15T 9株(有4株与katGS315T联合突变)。另有3株发生同义突变。37株Pto耐药的菌株中,8株发生点突变且均为C-15T,其中4株katGS315T和inhAC-15T双基因联合突变,余4株均为inhA单基因点突变C-15T。
aphC突变结果:标准株及5株全敏感MTB未发现aphC基因突变,233株异烟肼耐药菌中有1株菌株Leu106Val突变。
kasA突变结果:标准株及5株全敏感MTB未发现kasA基因突变,233株异烟肼耐药菌未有kasA基因突变。详见表3。
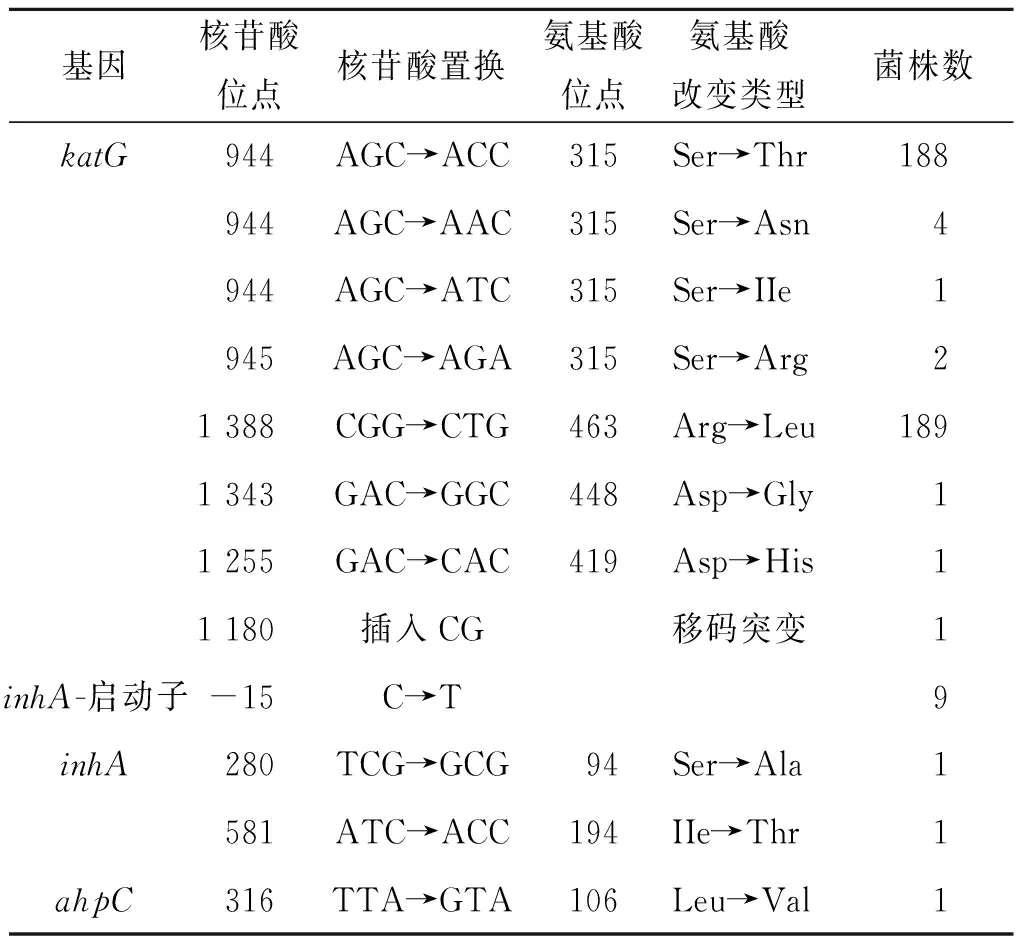
表3 233株异烟肼耐药菌株基因突变类型及位点情况Tab.3 Gene mutation types and sites in 233 isoniazid resistant strains
3 讨 论
异烟肼是结核病治疗的核心药物, 通过结核分枝杆菌过氧化氢酶过氧化物酶(katG编码)活化为杀菌形式。若katG发生突变,异烟肼活化效率降低或不能活化,导致结核分枝杆菌对异烟肼发生不同程度的耐药[6-7]。其他异烟肼耐药相关基因突变也参与异烟肼耐药,如inhA、ahpC、kasA等。其中,inhA是活化的异烟肼作用靶,inhA与kasA编码的蛋白合成酶参与分枝菌酸的生物合成,ahpC参与氧化-应激应答 (oxidativestress)[8]。
本项研究中,233株耐异烟肼菌株katG和inhA突变率分别为96.5%(223/233)、4.72%(11/233),二者的总突变率为98.7%(230/233)。katG第315位点突变是katG最常见的突变类型,本项研究katG315位点突变率为83.6%(195/233),与尼泊尔(81.4%)[9]及福建(86.7%)[10]相似,高于华东地区 (64.4%)[11]的报道,提示此位点突变发生率存在着区域性差异。据报道,katG315位点突变可能是由于katG氧化酶活性丧失且保留其过氧化氢酶活性,而这种基因的修饰具有生存优势,很容易在人群中传播,这也是其成为主要突变类型的原因之一[12-13]。本研究中katG315位点突变类型以S315T 188株最常见,其次为S315N 4株(2.1%),另S315R有2株、S315I仅1株。而katG463位点突变率(81.1%)虽较高,但据文献报道这主要是由于其所在的区域相对不稳定、极易发生改变所致,此位点的突变与INH耐药无关[14]。INH耐药菌株中inhA发生点突变率为4.72%(11/233),包括9株inhA启动子突变、2株inhA基因突变。inhA启动子突变以C-15T最为常见。国内外报道耐异烟肼菌株中,inhA基因突变所占比例从12%~60%不等[15-18],可见inhA基因突变存在着明显的地区性差异。
文献报道inhA基因突变常伴有katG突变,二者对INH有协同耐药作用[19],本研究中11株inhA基因突变中4株伴katGS315T突变,也证实这点。Seifert等[20]对2000-2013年的来自49个国家的118篇关于INH耐药基因突变的研究进行了系统综述,发现全球INH耐药菌株64%与katG基因315相关,19%由inhA基因-15位点突变所致。由于大多数MTB耐药发生于katG基因315和inhA基因-15位点,这两个位点已成为INH耐药基因检测的主要位点。本项研究中,katG315位点突变率83.6%,inhA-15位点的突变率4.72%,二者联合突变4株,检测此两个基因位点可检测出耐药基因突变率为85.8%(200/233),证实通过这两个位点的耐药基因型检测是可行的。
1株ahpC突变菌株对异烟肼耐药,可能是ahpC突变增强了AhpC的表达,降低过氧化物的浓度,进而抑阻了katG介导的异烟肼活化。233株异烟肼耐药菌未发现有kasA基因突变,与陈曦[6]、Mdluli[21]等国内外学者的研究结果不同,估计与扩增区域不同以及突变地区间差异有关,同时最近有报道kasA酶对INH耐药影响尚有争议[22-23],kasA基因突变与INH耐药的关系还需进一步研究。
Pto属碳硫胺类似药,作用机制与INH相似,需被MTB细胞质氧化还原酶作用成为细胞毒性成分,进而使MTB细胞膜不能合成霉菌酸及失去酸性,以此发挥抗结核功效[24]。两种药有共同的作用靶点:烟酰胺腺嘌呤二核苷酸(nicotinamide adenine dinucleotide,NAD),NADH依赖的烯酰-ACP还原酶(InhA,Rv1484)[25]。因此,inhA基因、inhA调控区域的突变,可致使inhA靶标过度表达或者修饰﹐降低与NAD之间亲和力而产生INH与Pto交叉耐药[23,26-27]。
本研究中,37株INH与Pto共同耐药菌株中,inhA发生点突变率为21.6%(8/37),8例均为inhA基因-15位点突变;虽有4例与katG基因联合突变,但Morlock等[26]研究表明未发现katG基因突变与Pto耐药相关。进一步证实Pto耐药与inhA基因-15位点突变密切相关,INH与Pto之间存在一定的交叉耐药率,且交叉耐药菌株中inhA基因-15位点突变最常见。虽有1例inhA启动子区域C-15T位点突变存在于INH耐药Pto敏感菌株,但可能与其他调节因子的代偿突变有关[28]。如研究所示,inhA基因-15位点突变在INH耐药株中所占比例较小,且传统药敏试验结果显示,INH与Pto的交叉耐药也只有15.8%。因此,INH耐药特别是无inhA突变的患者可选择Pto替代INH进行治疗。
综上所述,本研究发现重庆地区异烟肼耐药菌株基因突变率较高,大多数发生于katG基因315位点和inhA基因-15位点,通过检测这两个基因位点的分子耐药诊断学方法可以在重庆地区得到很好的应用,指导临床用药。同时INH与Pto存在着一定的交叉耐药,但耐药率较低,为INH耐药及耐多药结核病化疗方案中Pto的合理选择提供参考;Pto耐药的发生与inhA基因-15位点突变密切相关,这为inhA基因突变应用于INH与Pto交叉耐药快速诊断奠定了基础。
利益冲突:无
引用本文格式:朱大冕,刘文果,沈静,等. 重庆地区结核分枝杆菌异烟肼和丙硫异烟胺耐药及其交叉耐药相关基因突变研究[J].中国人兽共患病学报,2022,38(5):405-409. DOI:10.3969/j.issn.1002-2694.2022.00.047
