氮素改变辣椒初级代谢和薄壁组织厚度 调控比叶面积研究
2022-01-14LucasdeávilaSilva,RebecaP.Omena-Garcia,JorgeA.Condori-Apfata等
引言
比叶面积(SLA)是一个与叶结构相关的参数,通过测量单位干质量的投影叶面积进行量化,并与基于质量和面积的光合作用-氮关系的变化模式相 关[1]。SLA代表单位面积叶质量(LMA)的倒数[2], 且被认为是与在受控或自然环境下生长的一系列草本和木本物种的光合能力相关的重要生态指 标[3-6]。SLA[7-8]通过影响光捕获和CO2与RuBisCO 酶羧基化位点的结合进而影响光合能力[9-11]。该参数的变化源自叶片密度和厚度的变化。此外,叶片厚度[12-13]和LMA[2]因环境(营养、水分、光照状况等)和植物功能群的不同而不同,尽管LMA也可能独立于叶片厚度而发生变化[12]。有趣的是,LMA与栅栏和海绵状薄壁组织的厚度、上表皮表面和单位面积体积正相关,这进一步凸现了解剖结构对于全面理解LMA的重要性。同样,组织密度也会受到水分含量和细胞质成分的影响[14-17]。
许多环境因素,如光强和光质、UV-B光、CO2浓度、臭氧、土壤板结度、水分胁迫、温度、盐和养分有效性等,都会影响叶面积和叶质量之间的比率,从而影响SLA[2,18]。大量研究表明不同物种在多种环境下的叶密度和厚度可分别解释LMA差异的80%和20%[2]。在这种情况下,叶片特征的改变可归因于自然选择引起的种间差异,或环境条件下表型可塑性引起的种内变异[19]。相应的,那些能够根据资源供应而改变叶片形态、解剖结构和生理代谢的物种具有生态优势[20]。因此,SLA被视为物种之间以及不同环境之间的可塑性参数[21],从而SLA被广泛用于解释和理解不同物种生长模式的生态差异[2-4, 17, 22-23]。
一般来说,SLA与相对生长率(RGR)呈正相关,弱光下更明显。然而关于养分有效性对SLA的影响尚不清楚[3,21]。例如,干旱地区生长速率最高的桉树表现出较厚的叶组织,因为该地区没有光限制,从而表现出叶片较密和较厚的物种具有较高的水分利用效率[24]。在另一项关于非木本植物的研究中,在最佳营养供应和受控环境条件下种植时,种间RGR变异很大程度上受到SLA的影响[3]。在同一项研究中,自然条件下物种RGR和可用性氮之间的相关性分析表明,营养丰富环境中的自然选择有利于SLA较高的物种,而营养贫乏环境中的自然选择则相反[3]。因此认为,SLA与半干旱地区植物中的氮质量呈正相关,但在干旱地区SLA和氮质量无关,这表明SLA的科属依赖性反应可能对生态系统水平产生影响[23]。然而,这些在自然环境中进行的研究可能忽略了一个事实,即富氮环境可能与较高的植被覆盖率相关,从而降低了光的可用性,因此光是一个干扰因素。
养分中,土壤和叶片中的氮含量被认为是调节叶片形态和解剖结构的重要因素[18,23]。研究认为,随着叶片氮含量的增加,不同物种有较高的SLA(或较低的LMA)[2,12,25-30]。除氮素外,辐照度以及氮素和光照的复合因素也会影响SLA[31-33]。令人惊讶的是,氮供应对果实类植物,甚至是作物类植物的SLA影响仍未知。很少有在种内水平和作物品种之间探讨SLA的研究[8,26,30]。在这方面,我们之前已经证实:(1)施氮对C.chinense辣椒的生长和产量有较大影响,(2)C.chinense辣椒商品种Biquinho和Habanero在坐果率和果实大小、源库比和叶片形态方面存在显著差异[34]。因此,基于氮供应对C.chinense辣椒生长和果实产量的影响以及种内差异,研究分析了氮供应对SLA和相关性状的影响。本研究验证了在供氮和遮阴条件下两种C.chinense辣椒通过不同的代谢和结构来调节SLA的假设。首先,将两种基因型辣椒置于0~312.5 mg/dm3一系列氮素浓度下,研究了氮对SLA和初级代谢物的影响。其次,研究在解剖学水平上放大了氮效应,并研究了氮与其他常见的对SLA、叶片厚度和密度有影响的因素的交互作用。
1 材料与方法
1.1 植株生长条件
品种Biquinho和Habanero(TopSeed®, Agristar do Brasil Ltd.)均为Capsicum chinense辣椒,其种子在商品基质中发芽(Tropstrato®HT);45 d后,将幼苗移植到装有5 dm3黄-红土壤的花盆中,以确保试验结束时对根系发育没有限制。盆土经过筛选、混匀,并用CaCO3和MgCO3(摩尔比3.5∶1)调节至中性。之后,除氮外,土壤条件参照Novais 等[35]进行施肥,磷、钾、硫、硼、铜、铁、锰、钼和锌施肥总量分别为300、150、40、0.81、1.33、1.55、3.66、0.15和4.00 mg/dm3。由于土壤中的氮含量较低,NH4NO3以5 mg/dm3的浓度施入盆中,以确保移栽后植株的初始发育。为了避免土壤中的氮淋失,用塑料袋盖住盆底。以NH4NO3为氮源的第一次施氮是在第一分枝出现时进行的,这与生殖分化相对应。如前所述,该物候期是根据叶片和果实的生长速率及其对氮的需求来选择的[36-37]。试验过程中,每天供水,使土壤保持恒定的田间持水量。
为了评估氮剂量和遮阴对SLA的影响,连续两年进行独立试验。第1个试验中,从第一分枝出现 开 始,0、20、50、125和312.5 mg/dm3的 氮素,以溶液形式每20 d施于土壤中,总共5次等量补充[34]。第1个试验是在温室中进行的,室内温度为25℃ ± 5℃,平均辐照度为500 μmol/(m2·s)。植株花后(DAF)培养91 d。所有试验分析均在2016年7月—10月进行,在此期间平均光周期为11.5 h/12.5 h(白昼/夜间)(来源:美国华盛顿特区美国海军天文台天文应用部),平均辐照度为 1 300 kJ/m2(资料来源:巴西国家气象研究所)。第2个试验是为了分析氮供应和光照强度对SLA的影响。植株生长于温度为27℃ ± 5℃,平均辐照度为800 μmol/(m2·s)的温室当中。当第一分枝形成时,将一半的植株置于50%的遮阴下,而其他植株则保持在正常光强下。同时,每20 d施用20或125 mg/dm3的氮肥,以补充前述的4个等量[38]。试验分析均在2017年2月—4月进行,期间平均光周期为12 h 11 min / 11 h 49 min(昼/夜)(来源:美国海军天文台天文应用部),平均照度为1 390 kJ/m2(资料来源:巴西国家气象研究所)。植株花后培养74 d。
1.2 代谢物分析和氮含量测定
两个试验均在花后59 d 和30 d分别采集完全展开的第三片叶(从顶点往下数),采集叶片在液氮中快速冷冻,并储存在 - 80℃冰箱中等待后续研究分析。代谢物提取是先在液氮中研磨,然后称量质量,并用乙醇提取[39]。在乙醇提取物中,笔者测定了蔗糖、果糖、葡萄糖[40]、苹果酸[41]、总氨基酸[39]、 NH4
+[42]、NO3-[43]和叶绿素a、b的含量[44]。淀粉和蛋白含量按Bradford[45]和Fernie等[40]所述,用乙醇不溶性部分测定。总氮含量则参考Kalra (1998)[46]的方法于每次试验结束时采集叶片进行测定。
1.3 比叶面积、解剖特征和叶密度的测定
参考Cavate等[47]的方法,两个试验的SLA均使用叶盘测定。第1个试验中,从花后88 d植株的成熟叶片上采集9个1.03 cm2的圆盘状叶片。第2个试验中,在花后10、30、50和70 d时采样,每个处理分别采集6个圆盘状叶片。研究所用结果为平均值,因为试验发现随着时间的推移处理间没有差异。此外,研究采用了第2种方法(Poorter和Remkes等[3]提出)测定SLA,即叶面积与其干质量之比(m2/kg)。在该方法中,在花后72 d使用叶面积仪(Li 3100C Li Cor,Inc.,Lincoln,NE,USA)测量每处理植株的总叶面积。之后,在80℃下对叶片进行烘干称量质量,然后计算SLA。值得注意的是,这两种方法的结果相似(附表S1)。
花后70 d(第2个试验)进行叶片解剖结构分析。为此,采集第三分枝上处于中等光照强度的完全展开的健康叶片,在FAA50[48]中真空固定48 h,保存在70%乙醇中。根据使用说明,每处理随机选择3个样品,使用乙醇进行梯度脱水,并用甲基丙烯酸乙二醇酯(徕卡®包埋试剂盒)包埋处理。在旋转切片机(型号RM2155,美国伊利诺伊州迪尔菲尔德莱卡微系统公司)中制作8 μm厚的横截面,用甲苯胺蓝[49]染色10 min后,用水冲洗并在室温下干燥。载玻片用合成树脂固定(Permount,Fisher),以 便 在 光 学 显 微 镜(AX70-Olympus)中进行观察和记录。利用Image Pro Plus软件(MediaCybernetics)对显微照片的叶片进行化学计量分析。因此,试验对每处理的3株9个区域进行了全叶厚度、栅栏薄壁组织厚度、海绵薄壁组织厚度和细胞间隙百分比的测定。根据第2个试验,根据以下方程式计算叶片密度:
叶片密度=圆盘状叶干质量/(圆盘状叶面积×叶横截面直径)
根据第2个试验的描述进行叶盘取样,测定叶盘在已知面积内的干质量(打孔器面积为1.03 cm2)。上述圆盘叶来自于处于同一个体发育时期和生理阶段的叶片,这些叶片均为完全展开、成熟、健康和活跃的叶片。根据叶片横截面测量叶片宽度。
1.4 统计分析
第1个试验采用完全随机区组设计,变量为基因型和供氮量,共有5个重复。数据的正态性通过夏皮洛-威尔克法检验矫正,然后对所有变量进行双因素方差分析。使用多重比较法进行基因型差异显著性分析(P≤ 0.05),根据供氮量进行回归模型参数调整。第2个试验采用裂区设计[50],3个变量:遮阴(光照和遮阴)为主处理,氮供应(20或125 mg/dm3)为副处理,两个基因型(Biquinho和Habanero)为子处理。每处理有6个生物学重复。通过夏皮洛-威尔克法检验数据的正态性,然后进行三因素方差分析。采用F检验和多重比较法检验实现各因素差异显著性的分析(P≤ 0.05)。对于特定叶面积,第1个试验中每处理5株重复,第2个试验每处理6株重复。叶片解剖和计量统计学分析每处理3个生物学重复。每株有3个组织学重复,因此每处理有9个切片。使用SigmaPlot v.11.0(美国加利福尼亚州圣何塞市Systat软件公司)和R语言[51](R核心团队2019)软件绘制图表,并进行均值比较、回归分析和皮尔逊相关分析。
2 结果
2.1 氮素供应对SLA的影响
为了明确供氮量对辣椒(C.chinense)SLA的影响,本研究在5个氮剂量下,种植了在坐果率、果实大小、库源比和叶片形态方面存在差异的两个商品种(Biquinho和Habanero)[34]。每个品种以供氮量为变量调整回归模型从而阐明SLA的变化(图1a)。由图1可知,不同处理下,两品种的SLA均随氮供应的增加而下降。在20 mg/dm3和50 mg/dm3的氮素处理下,品种Biquinho的SLA高于品种Habanero(图1a)。
在第2个试验中,结合两个辐照度水平(800和400 μmol/(m2·s),研究两种基因型的辣椒在中度缺氮和充足氮(20和125 mg/dm3)条件下SLA的变化。由表1可知,光照、供氮和基因型对辣椒SLA有显著的三重交互作用(P= 0.041)。因此,可以根据基因型,确定光照或氮供应水平之间是否存在差异(图1b)。两种基因型叶片的SLA在遮阴条件下都有所增加,但只有Biquinho的SLA在供氮量变化时表现出显著差异(图1b)。值得注意的是,尽管第1次500 μmol/(m2·s)和第2次800 μmol/(m2·s)试验的温室辐照度不同,两个品种在不同供氮量下的SLA表现出相似的变化(图1)。
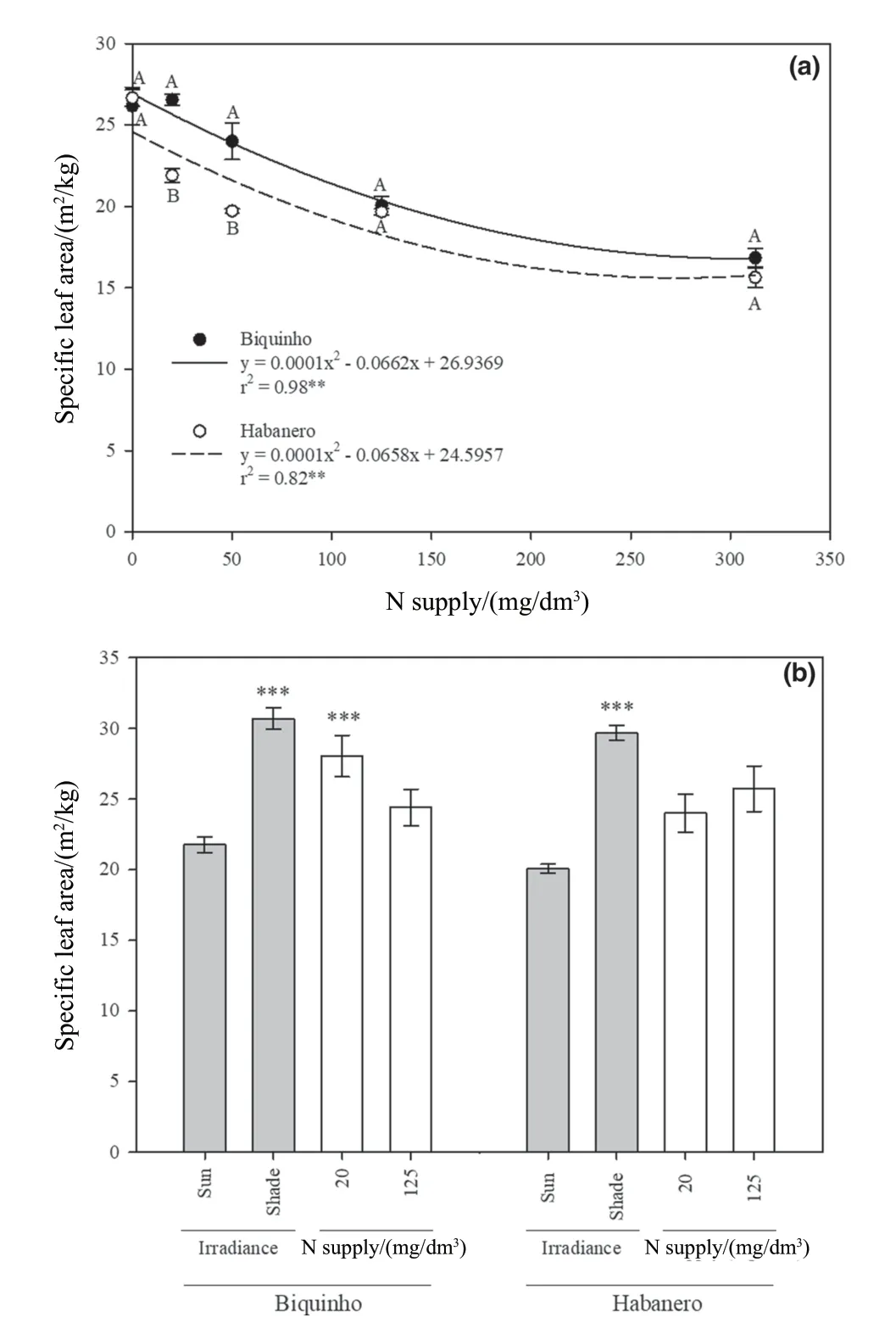
图1 a.5个氮水平和500 μmol/(m2·s)处理下,花后88 d两个辣椒品种的SLA; b.光照水平800和400 μmol/(m2·s)、氮供应(20和125 mg/dm3)和基因型(Biquinho和Habanero)对SLA的三重影响
研究每株植物的总叶面积除以单株叶片的总干质量以便进一步明确SLA的变化。正如预期的那样,该分析证实了遮阴可提高两种辣椒的SLA。在光照和遮阴条件下,SLA的值由19.36 ± 0.75 m2/kg 变为27.49 ± 0.88 m2/kg(表1和附表S2)。此外,由表1可知针对SLA,氮供应和基因型存在互作(表1)。结果表明,在20 mg/dm3氮条件下,品种Biquinho叶片表现出较高的SLA,而Habanero的SLA未观察到氮效应(图2a)。
2.2 供氮对不同基因型叶片和薄壁组织厚度影响的差异
接下来,研究决定评估氮供应和辐照度水平对SLA相关参数的影响。遮阴对相关解剖性状的影响是独立于供氮或基因型之外的,而供氮效应与基因型有关(表1)。Habanero的植株在20 mg/dm3的氮处理下表现出较厚的叶子,而Biquinho的叶片厚度没有影响(图2b)。但在125 mg/dm3的氮处理下,Biquinho表现出较厚的栅栏组织,而对Habanero的栅栏薄壁组织厚度没有受到氮供应的影响(图2c,3)。相比之下,在20 mg/dm3的氮处理下,Habanero表现出更厚的海绵状薄壁组织,而Biquinho的该组织厚度与氮供应无关(图2 d,3)。此外,在20 mg/dm3的氮处理下,Habanero比Biquinho的海绵状薄壁组织更厚,而在125 mg/dm3的氮处理下两个材料无明显差别。总的来说,这些结果表明Biquinho主要表现为栅栏薄壁组织的改变,而Habanero显示了海绵状薄壁组织的变化,以响应不同的氮供应(图3)。

图2 对氮供应(20或125 mg/dm3)和C.chinense(Biquinho或Habanero)基因型交互作用下的(a)比叶面积(SLA)(花后74 d的总叶面积与叶干质量比)、(b)叶厚度、(C)栅栏和(D)海绵状薄壁组织(平均值± SE)进行方差分析(表1)

图3 花后70 d,遮阴和光照下,中度缺氮(20 mg/dm3)和氮充足(125 mg/dm3)时Biquinho和 Habanero两个品种成熟叶片的横截面
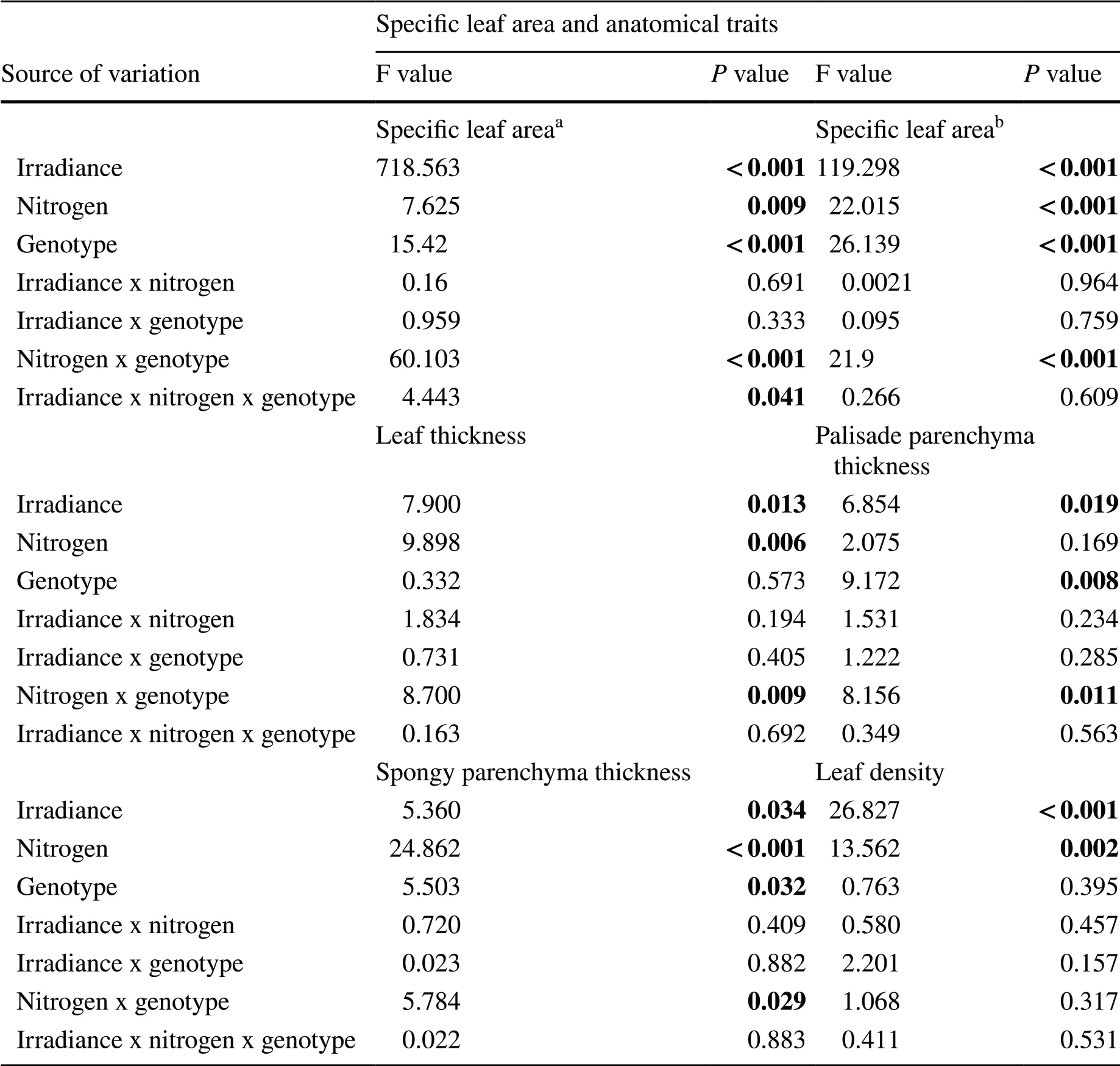
表1 利用三因素方差分析法以表明光照(光照和遮阴)、供氮(20或125 mg/dm3)、C.chinense基因型(Biquinho或Habanero)及其互作对成熟叶片的比叶面积、叶面厚度、栅栏和海绵状薄壁组织的影响(第2个试验)
在第2个试验中,SLA分析是指开花后10 d、30 d、50 d、70 d(它们之间没有差异)[47]和74 d[3]收获的叶盘平均数表示,同时选用开花70 d的材料进行解剖特征和叶密度的测定。数据用F值表示,且通过P值进行显著性分析。粗体数值表示数据通过F测验在P≤ 0.05下的统计学意义。
aSLA的测定参照Cavate等[47]的方法。
bSLA的测定通过总叶面积除以叶片的总干 质量[3]。
此外,我们观察到,遮阴对解剖特征的影响与氮供应或基因型无关(附表S2)。与光下叶片厚度(217.7 ± 8.5 μm)相比,遮阴下两个品种的叶片厚度较薄(191.6 ± 8.7 μm)(图4a)。这与遮阴条件下较薄的栅栏和海绵状薄壁组织的结论一致(图4b,c)。遮阴和光照下栅栏薄壁组织的厚度分别为69.9 ± 2.6 μm和80.8 ± 4.8 μm(图4b),海绵状薄壁组织厚度分别为92.9 ± 6.4 μm和105.9 ±5.9 μm(图4c)。
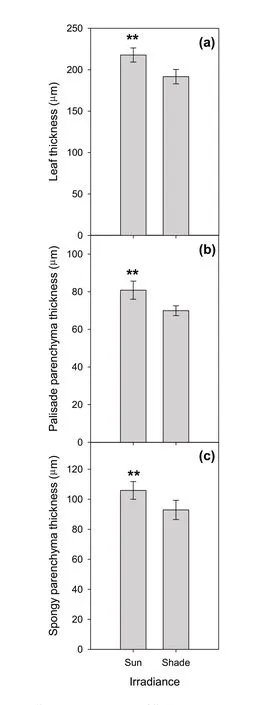
图4 花后70 d,光照对两种辣椒(Biquinho和Habanero)叶片的厚度(a)、栅栏组织厚度(b)和海绵组织厚度(c)的影响
与解剖特征相反,基因型对叶片密度没有显著影响(表1)。遮阴条件下Biquinho和Habanero的叶片密度较光照下的低(图5)。随着氮肥用量的增加,两种辣椒均表现出较高的叶片密度(图5)。
2.3 氮含量和氮代谢产物与SLA相关
为了确定氮供应对碳氮相关的代谢物水平的影响,根据第1个试验的氮供应分析了初级代谢物的变化(附图S1)。接下来,对SLA值、氮含量和碳氮相关代谢物之间进行皮尔逊相关分析(图6)。皮尔逊相关分析是在两个品种的代谢物、叶片性状以及碳氮含量等与SLA正相关或负相关全部参数的基础上进行的。氮含量与氮相关代谢物如氨基酸、蛋白质、NH4t">+和NO3-之间呈正相关。另一方面,氮含量及氮相关代谢物含量都与SLA呈负相关(图6)。相反,碳相关代谢物如淀粉、葡萄糖和苹果酸的含量与SLA呈正相关 (图6)。
为了在第2个试验中验证初级代谢物对SLA的影响,利用之前对C.chinense[34]的研究获得的数据,对不同光照及氮水平下对SLA和氮含量、氨基酸、蛋白质和淀粉含量进行皮尔逊相关分析。在20~125 mg/dm3氮范围内,品种Biquinho的SLA随着施氮量的增加(以及氮代谢产物)和栅栏薄壁组织厚度的增加而降低(图2,7)。另一方面,在品种Habanero中观察到的SLA微小变化,可能是为了响应氮供应增加,氮相关代谢物含量的轻微增加和叶片厚度(海绵状薄壁组织)降低相互抵消后的结果。
3 讨论
本次研究比较了在坐果率、果实大小和叶片形态上具有差异的两个C.chinense种辣椒(Biquinho和Habanero),以便进一步了解氮、光强及其互作对果实类植物SLA的影响。我们之前的研究表明,辣椒植株叶片中的氮含量与施氮量成比例增加[34]。本研究表明,SLA与品种无关,各品种SLA均随N剂量的增加而降低(图1a)。为了更好地理解这一效应,研究分析了不同光照水平以及在氮缺乏和充足情况下影响叶片密度、厚度及调节SLA因子[52]的性状。遮阴条件下两个品种的SLA均有所增加,且基因型和N剂量之间存在交互作用(图1)。叶片密度随着较高的细胞含量(如细胞内含物、淀粉粒和晶体)和较高的细胞壁沉积[13,52]或更小的空气空间体积而增加[2]。现有的研究中,在叶片横截面上,栅栏薄壁组织比海绵薄壁组织显示出更少的细胞间隙(图3)。这两个品种在叶片结构参数(如栅栏和海绵状薄壁组织)方面对氮肥的反应不同(图2)。Biquinho的栅栏薄壁组织厚度增加(图2c),而Habanero的海绵薄壁组织减少(图2d)。有趣的是,Habanero的叶片厚度减小,而Biquinho在较高氮剂量下却保持稳定(图2b)。上述结果表明,这两个品种具有明显的叶肉细胞扩张,这可能会调节其SLA和叶片厚度,但保持稳定的叶片密度(图5)。

图5 花后70 d,不同光照水平800和400 μmol/(m2·s)与N供应(20和125 mg/dm3)水平下Biquinho和Habanero辣椒的叶片密度
考虑到叶密度与代谢产物的关系,所有非结构性碳水化合物(可溶性糖和淀粉)与蛋白质的总和可能对叶生物量有相当大的影响[2,53-55]。本研究中,所有初级代谢产物贡献了叶片干质量的9.5%~28.1%(附图S1),以响应氮供应量的增加。与其他代谢物相比,氨基酸、蛋白质和淀粉含量的变化幅度更大。氨基酸在干叶中的质量为0.7%~17%,蛋白质占干叶质量的0.6%~3.8%,两者均与氮含量正相关,与SLA负相关(图6)。另一方面,淀粉(占干叶质量的2.1%~10.3%)与氮含量负相关,与SLA正相关(图6)。与之一致的是,前人的研究表明,缺氮会导致碳代谢改变,例如苹果酸和其他有机酸水平降低,淀粉水平增加[56-57]。 总之,当氮供应增加时与氮代谢相关的代谢物的合成是一个重要特征,此特征也与叶片密度增加有关(图5)。

图6 5个氮水平下,Biquinho和Habanero两个辣椒品种的SLA、叶特性、碳含量、氮含量及碳氮相关代谢物间的皮尔逊相关分析
Poorter等[2]认为在与氮代谢相关的细胞成分中,蛋白质水平可能对SLA产生影响。蛋白质含量的数量变化可能超过叶子中非结构碳水化合物的变化[58]。本研究的结果表明氨基酸水平的变化也是决定SLA响应氮供应变化的重要因素(图7和附图S1)。某些氨基酸水平有利于调节硝酸盐的吸收和氮同化[59-60],同时也有助于渗透调节[61],而渗透调节反过来又在细胞伸展中发挥有效作用[62]。因此,除了代谢作用外,氨基酸可能还具有结构功能,不仅通过参与增加叶片密度直接影响SLA,而且可通过调控叶片解剖结构来影响SLA。总的来说,由养分供应驱动的叶面积和叶质量之间关系的变化与叶密度的变化比与单位面积叶体积的变化关系更密切[2,8]。因此,我们推测两种基因型(图1a)的SLA反应,至少部分的可以用氮和碳代谢相关初级代谢物(附图S1)来解释(图5)。然而,还需要进一步的研究来充分阐明这些代谢物在控制辣椒叶片密度方面的具体功能。
由于两种基因型在氮和碳相关代谢物对叶片密度等性状影响上表现出相似的行为(附图S1),因此研究认为氮由中度缺乏到充足供应(图1b、2a)时,基因型之间的SLA不同源于叶片解剖结构的可塑性不同(图2b-d)。氮充足时,由于叶片密度的增加和栅栏薄壁组织的增厚,品种Biquinho的SLA降低(图2c、3、5)。相反,随着氮供应的增加,品种Habanero由于海绵薄壁组织的变化导致叶片厚度的下降[30],厚度变薄补偿了叶片密度的变化,从而使SLA没有变化(图2b、d、3、5)。此外,由于栅栏薄壁组织比海绵薄壁组织密度更大[63],在中度缺氮至充足氮范围内,品种Biquinho的SLA敏感性更高。Xiong等[8]研究不同品种水稻叶面积/叶质量比对施氮变化的响应差异时也发现了类似的现象。
本研究明确了Biquinho和Habanero两种基因型辣椒SLA变化的不同原因。研究结果表明,两种基因型辣椒在氮供应量增加时SLA均有所降低,这是因为氮和碳相关代谢物(主要是氨基酸、蛋白质和淀粉)的转移导致叶片密度增加(图7和附图S1)。然而,在中度缺氮到氮充足范围内,辣椒的SLA也受到材料表型可塑性的影响。随着供氮量的增加,SLA在两个基因型中表现出不同的变化,品种Biquinho具有更高的叶密度和更厚的栅栏薄壁组织,而Habanero辣椒则观察到更高的叶密度和更薄的叶片厚度(与更薄的海绵薄壁组织 有关)。
虽然本研究没有发现细胞间隙比例的改变,但研究氮供应对叶细胞数量、细胞壁厚度、细胞扩张将有助于在分子水平上更好地理解氮对SLA及器官发生的影响[64]。了解环境条件如何影响叶片形态对于固定栖息地中的物种生存非常重要[31]。与其他物种类似[3,31-33],Biquinho和Habanero采用了类似的策略来降低SLA以保证在弱光环境下存活。虽然两个品种在极低和极高氮剂量下表现出相同的行为,但在中等氮剂量下栽培时表现不同。由此,在极端氮水平下Biquinho和Habanero辣椒在代谢方面表现出和其他研究一样的响应行为。然而,在中度缺氮或氮充足的条件下,两种辣椒采用不同策略与结构特征去调节SLA。推测这些品种的自然(或繁殖)栖息地中的氮可用性可以显著影响SLA行 为[3]。鉴于SLA是决定不同物种在其自身环境中适应性的一个重要特征,因此有必要进一步研究SLA对氮供应的分子、结构和生理调节响应。
作者贡献
LAS, JAC-A和AN-N设计试验。LAS和JAC- A执行试验和采样。LAS, JAC-A, PMAC, EATP和NMS进行分析测试。LAS, RPO-G, EATP和AN-N撰写论文。FMD, RPO-G, PMAC, AZ, WLA, RS, EATP和AN-N审阅修订论文。
Acknowledgements
由国家科学技术发展委员会资助(CNPq)(ANN 306818/2016-7号,LAS 501090/2015-0号,WLA 402511/2016-6号)。米纳斯吉拉斯州研究扩展基金会(FAPEMIG)[授权编号:CRARED-00053-16 ANN]。感谢CNPq向RPO-G(流程号152121/2019-6)、ANN、FMD和WLA授予的研究经费。RS得到了爱尔兰农业、食品和海洋部(DAFM)研究经费(VICCI编号:14/S/819)的支持。我们还感谢Franciele Santos Oliveira女士对样品制备的支持。
