针刺对大鼠运动性骨骼肌损伤内质网应激的干预作用及机制*
2021-10-22丁海丽黄增浩任在方孙竹昕李俊平王瑞元
丁海丽, 黄增浩, 任在方, 孙竹昕, 李俊平, 王瑞元
(1. 成都体育学院运动医学与健康学院、运动医学与健康研究所, 四川 成都 610041; 2. 北京体育大学运动人体科学学院, 北京 100084; 3. 中日友好医院, 北京 100029; 4. 清华大学继续教育学院, 北京 100080)
骨骼肌是运动的直接效应器官,大负荷运动尤其是不习惯的离心运动,会使肌纤维超微结构发生改变,呈现出肌肉僵硬、肌力下降以及主观感觉疼痛等症状和体征,称为运动性骨骼肌损伤(exercise-induced muscle damage,EIMD),可引起延迟性肌肉酸痛(delayed onset muscle soreness, DOMS)的亚临床疼痛现象。EIMD及DOMS若不及时治疗,会加剧运动损伤,影响运动表现(exercise performance)以及锻炼者正常生活[1-3]。EIMD一直以来都是运动医学和运动生理学研究的热点问题,寻找有效干预手段来恢复或缓解EIMD进而减轻DOMS症状,对提高运动成绩和帮助锻炼者重返运动场具有重要意义。
针灸作为中医传统疗法之一,治疗骨骼肌损伤历史悠久、疗效确切。研究团队早期由卢鼎厚教授伊始[4]按照《灵枢》中“以痛为腧”治疗经筋病的记载,采用“阿是穴-斜刺”干预EIMD,效果显著。随后,团队王瑞元等对EIMD和DOMS发生的机制以及针刺干预效应与作用靶点进行了大量实验研究,包括骨骼肌收缩蛋白代谢、泛素蛋白酶体途径[5]、生肌因子表达[6]、线粒体动力学[7]以及钙离子信号等。课题组前期研究[8]证实了运动后针刺可改善骨骼肌内质网功能受损,逐渐恢复内质网高钙浓度且有效降低内质网应激(endoplasmic reticulum stress, ERS)程度,初步探讨了Ca2+调节机制与蛋白折叠的协同效应。为了进一步探讨针刺干预EIMD内质网途径的相关机制,本实验从内质网功能酶学指标、内质网应激和PERK通路三方面,综合分析针刺治疗运动性骨骼肌损伤的内质网途径机制与靶点效应,以期为传统针刺手段介入运动损伤的治疗和恢复提供新的实验依据与理论支撑。
1 材料与方法
1.1 实验动物与分组
健康雄性8周龄Spraque Dawley(SD)大鼠96只,SPF级,体重(225.77±11.03)g,使用许可证号:SCXK(京)2012-0001,由北京维通利华实验动物技术有限公司提供。在北京体育大学科研中心实验动物房饲养和训练,自由饮食饮水,室内温度20~26℃,相对湿度40%~70%,12 h明暗交替。适应性喂养3 d后,按随机数字表法分为4组,即空白对照组(control,C)、单纯运动组(exercise,E)、针刺对照组(acupuncture,A)和运动针刺组(exercise+acupuncture,EA)。E组、A组和EA组根据运动、针刺干预后不同时相,又分为0 h/12 h/24 h/48 h/72 h各5个亚组,共16个组别,每组6只。
1.2 运动造模方案
参照Armstrong等[9]的离心运动方案,通过一次大负荷运动建立EIMD模型。使用小动物电动跑台训练, E组和EA组先进行3 d适应性训练,适应性训练后开始正式实验,坡度-16°,速度16 m/min,时间90 min。
1.3 针刺治疗方法
运动后即刻对EA和A组大鼠施以针刺干预。固定器固定大鼠,消毒双后肢,用直径0.25 mm的针灸针[汉医牌,2009第2270002号]于大鼠小腿跟腱上0.5 cm处沿小腿三头肌纵向,从远端斜刺穿过肌腹,进针角度约30°,留针2 min。课题组前期研究中,通过小腿三头肌局部解剖学手段发现,此方案可复制损伤肌肉——比目鱼肌(长时间运动主要动员的慢肌之一)的模型。
1.4 取材与样本采集
在各组对应时相将大鼠称重后腹腔麻醉,按 3.5 ml/kg体重注射10%水合氯醛),经腹主动脉取血后,迅速分离比目鱼肌。冰冷生理盐水漂洗,除去血液,滤纸拭干,剔除结缔组织。左侧取2 mm×3 mm组织块投入2.5%戊二醛固定液,制备电镜样品,剩余组织剪碎后组织匀浆,待测ELISA实验;右侧组织置于液氮迅速冷冻后-80℃冰箱保存,待测Western blot实验。
1.5 指标检测
1.5.1 透射电镜观察肌纤维超微结构:取C组、A组、E组和EA组大鼠制备电镜样品,经戊二醛固定,锇酸再固定,丙酮溶液脱水,环氧树脂Spurr包埋后,制备超薄切片,醋酸双氧铀和柠檬酸铅双染色。采用H-600IV型透射电子显微镜观察骨骼肌纤维超微结构。
1.5.2 酶联免疫吸附(ELISA)法测定SERCA、PDI含量:在酶标板空白孔中加入100 μl样品,空白对照加入100 μl蒸馏水,各孔加入50 μl酶标记溶液,密封后37℃孵育1 h;洗涤并拍干,加显色剂A、B各50 μl,室温避光10 min;加50 μl终止液,置于酶标仪30℃孵育2 h;在550 nm波长处每隔1 min连续测定各孔光密度OD值。
1.5.3 蛋白质免疫印迹(Western blot)法检测GRP78、p-PERK和p-eIF2α表达:BCA蛋白定量试剂盒测定蛋白浓度,配制12%分离胶,5%浓缩胶,上样量20 μg/well;浓缩胶恒压90 V,约20 min,分离胶恒压120 V。湿转法转膜,丽春红染色,观察转膜效果。封闭PVDF膜。5%BSA-TBST稀释一抗(1∶1 000),5%BSA-TBST稀释HRP标记山羊抗兔IgG(1∶10 000)。胶片曝光,显影,定影。使用图像分析软件Image J检测各条带的灰度值,以GAPDH为内参,将目的蛋白条带灰度值和GAPDH条带灰度值的比值表示蛋白的表达水平。
1.6 统计学处理
2 结果
2.1 针刺对各组大鼠肌纤维超微结构的影响
C组肌节清晰,A带与I 带分界清楚、排列规则,Z 线整齐且两侧线粒体均匀分布。A组各时相肌小节、Z线以及线粒体排列与C组相近,无明显变化。E组0 h~48 h出现不同程度的Z 线加宽、断裂,明暗带分界不清,线粒体肿胀并聚集在肌膜下,72 h虽有所缓解,但仍未完全恢复。EA组对应时相比E组超微结构均明显改善,在72 h肌节结构整齐,未见Z 线和Z 线断裂,线粒体在肌膜两侧排列整齐(图1)。
2.2 针刺对各组大鼠肌组织SERCA含量的影响
与C相比,A组各时相SERCA含量均无显著变化(P>0.05);E组SERCA含量呈降低后逐渐回升趋势、在0 h、12 h、24 h和48 h均显著降低(P< 0.05);EA组SERCA含量虽有所降低但48 h已无显著差异(P>0.05)。与E组对应时相相比,EA组SERCA含量在48 h和72 h显著升高(表1)。

Fig. 1 Ultrastructural changes in different phases in each group (×8 000)
2.3 针刺对各组大鼠肌组织PDI含量的影响
与C相比,A组各时相PDI含量均无显著变化(P>0.05);E组各时相PDI含量亦无显著变化(P>0.05);EA组PDI含量各时相则显著升高(P< 0.05)。与E组对应时相相比, EA组PDI含量0 h至72 h各时相均显著升高(P<0.05,表2)。
2.4 针刺对各组大鼠肌组织GRP78、p-PERK、p-eIF2α表达的影响
与C组相比, A组GRP78表达各时相均无显著变化(P>0.05);E组GRP78表达在运动后均处于较高水平,0 h、12 h、24 h、48 h和72 h均显著升高(P<0.05);EA组仅在0 h显著升高(P<0.05)。与E组对应时相相比,GRP78表达均显著降低(P<0.05,表3,图2)

Fig. 2 Comparison of GRP78 protein expression in different phases of each group
2.5 针刺对各组大鼠肌组织p-PERK和p-eIF2α表达的影响
与C组相比,A 组p-PERK和p-eIF2α表达各时相均无显著变化(P>0.05);E组p-PERK和p-eIF2α表达在运动后0 h、12 h和24 h显著升高(P<0.05);EA组仅在0 h显著升高(P<0.05)。与E组对应时相相比,p-PERK和p-eIF2α表达12 h和24 h显著降低(P<0.05,表4,图3,表5,图4)
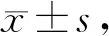
Tab. 1 Comparison of SERCA content in different phases of each group (ng/ml, n=6)
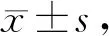
Tab. 2 Comparison of PDI content in different phases of each group (ng/ml, n=6)
Tab. 3 Comparison of GRP78 protein expression in different phases of each group(% C group, n=6)
Tab. 4 Comparison of p-PERK protein expression in different phases of each group(% C group, n=6)
Tab. 5 Comparison of p-eIF2α protein expression in different phases of each group(% C group, n=6)
Fig. 3 Comparison of p-PERK protein expression in different phases of each group
Fig. 4 Comparison of p-eIF2α protein expression in different phases of each group
3 讨论
骨骼肌细胞内质网是蛋白质加工和钙离子贮存的主要场所,对应激极为敏感,耗竭内质网Ca2+、二硫键错配、突变蛋白质表达等均可诱发ERS,运动应激能够引起骨骼肌内质网 Ca2+通道及相关酶功能改变,且与EIMD的发生密切相关[10-13]。目前关于针刺干预EIMD内质网作用途径的研究主要集中Ca2+调节方面,而对其另一功能——蛋白折叠修饰,则涉及较少。因此,本研究着重观察针刺对运动诱导骨骼肌损伤大鼠内质网Ca2+-ATP酶(sarcoplasmic reticulum Ca2+-ATPase, SERCA)、蛋白二硫键异构酶(protein disulphide isomerase, PDI)、内质网应激标志-葡萄糖调节蛋白78(glucose-regulated protein 78, GRP78)和通路蛋白PERK的影响,从内质网途径探究针刺治疗运动性骨骼肌损伤的作用机制。
本实验观察到针刺可缓解EIMD肌纤维结构改变,改善肌小节紊乱、线粒体肿胀和肌膜下聚集等。在针刺影响EIMD大鼠内质网钙泵功能方面,运动针刺组的SERCA含量在48 h和72 h显著高于单纯运动组,提示针刺上调大负荷运动后SERCA含量方面,并没有在早期出现,而是48 h或72 h较明显。推测针刺可以缓解内质网功能受损,有利于内质网回收钙及恢复高钙浓度,但SERCA可能并未及时发挥针刺的靶点作用。基于此,本研究对针刺干预后内质网折叠功能的二硫键异构酶PDI也进行测定,PDI具有自主异构酶活性,并在错误折叠蛋白聚集时增多,通过水解二硫键而发挥重要作用[14]。结果表明,单纯运动组PDI含量仅在运动后即刻显著高于对照组水平,运动针刺组PDI含量在干预后即刻、12 h、24 h、48 h和72 h均明显高于运动组。提示运动后进行针刺干预,可明显提升骨骼肌PDI含量,从而有效应对内质网应激的未折叠、错误折叠蛋白聚集,使其正确折叠和组装,相对于SERCA的变化情况,PDI的反应较为及时和快速,该结果提示,PDI可能具有一定靶点作用,但其确切作用途径和分子机制还有待进一步探究。
当生物组织细胞处于缺氧、氧化应激等状态时,细胞亚单位内质网会在一系列信号级联反应下产生应激反应,诱发ERS和未折叠蛋白反应,适度ERS可应对细胞环境扰动并恢复内质网良好的蛋白折叠环境,过度ERS则引起细胞功能失调、自噬或凋亡[15-17]。诱导GRP78生成是内质网应激的一个标志特征,非应激状态下,伴侣分子GRP78位于内质网腔,与蛋白激酶R样内质网激酶(protein kinase R-like ER kinase, PERK)、肌醇酶1(inositol requiring enzyme 1, IRE1)和活化转录因子6(the activating transcription factor, ATF6)三个ERS感受蛋白结合,处于非活化状态[18]。发生ERS时,GRP78与三个感受器解离从而激活三条信号通路,引发一系列信号转导反应,当应激时间过长或强度较大,超出机体自我调整能力,即可能引发细胞损伤和不良结局。PERK可感知内质网中Ca2+水平,通过磷酸化真核细胞起始因子(eukaryoticinitialion faclor, eIF2α)抑制蛋白合成,mRNA翻译能力下降,进而活化转录因子4(the activating transcription factor, ATF4)后上调C/EBP同源蛋白(C/EBP homologous protein, CHOP)基因表达,参与细胞的整合应激反应过程。目前,已有不少国内外学者对针刺影响PERK通路蛋白的效应机制进行了报道。Zhang等研究表明[19],冬眠松鼠骨骼肌ERS过度引发的细胞凋亡可以通过PERK信号通路介导的p-eIF2α和GRP78上调而缓解。郁洁[20]等研究发现,针刺可以下调实验性脑缺血大鼠内质网应激PERK通路重要因子eIF2α的基因表达,缓解脑缺血性内质网应激状态。本实验中GRP78的变化表明,针刺可缓解EIMD大鼠骨骼肌ERS,对PERK通路蛋白的检测结果表明,针刺可明显下调运动后12 h和24 h p-PERK与p-eIF2α表达,提示PERK具有一定的靶点作用,而对于另外两条通路IRE1和ATF6的变化趋势尚有待进一步探讨。
综上,针刺可有效缓解EIMD大鼠骨骼肌超微结构受损情况,其作用机制可能与上调PDI及抑制内质网应激PERK通路相关。
