基于LOX-HPL 途径的武夷肉桂加工中香气物质的形成与调控
2021-03-18周子维刘宝顺武清扬毕婉君倪子鑫赖钟雄
周子维 , 刘宝顺 , 武清扬 , 毕婉君 , 倪子鑫 , 赖钟雄 , 孙 云
(1.福建农林大学 园艺学院/茶学福建省高校重点实验室,福建 福州 350002;2.福建农林大学 园艺植物生物工程研究所,福建 福州 350002;3.武夷山市幔亭岩茶研究所,福建 武夷山 354300)
武夷岩茶是我国的传统名茶,它是在武夷山自然生态环境条件下,选用适宜茶树品种的鲜叶为原料,经晒青、做青、杀青、揉捻、干燥、拣剔、筛分、匀堆、焙火等工艺制成的乌龙茶,具有“岩骨花香”的品质特征[1]。 作为武夷岩茶的主要花色品种,武夷肉桂因香气馥郁,似有辛辣的桂皮香而得名[2-3],颇受市场青睐。
香气是衡量茶叶品质的重要指标之一,除茶树品种所赋予的基底外,更多源自茶树鲜叶采摘后的加工工艺[4-6]。 目前,茶树鲜叶中已被分离鉴定的挥发性物质约有80 余种, 乌龙茶中约有近400 种[7]。脂肪族类挥发性成分是茶叶中重要的香气组分之一[8],茶树鲜叶在采后的萎凋、做青过程中产生的高温、机械损伤等非生物胁迫,诱导了不饱和脂肪酸的脂氧合酶代谢途径(LOX pathway)[1]中结构基因的表达,进而提升关键酶活力,促使离体鲜叶中以亚油酸、α-亚麻酸为代表的不饱和脂肪酸氧化降解,形成丰富的脂肪族类香气物质[9-10]。 根据LOX pathway 中关键限速酶LOX 的氧化产物氢过氧化物(9/13-HPOT) 的代谢差异, 可分为 LOX-AOS 和LOX-HPL 两个支路途径[11-12], 前者在逆境胁迫中13S-HPOT 在丙二烯合酶(AOS) 、丙二烯氧化物环化酶(AOC)作用下形成12-氧植物二烯酸(OPDA),该物质再历经还原和3 次β-氧化反应后形成茉莉酸(JA),这是植物在生长发育过程中,最为迅捷的应激信号物质之一[13]。 后者主要响应外源生物及非生物胁迫,在 HPL、ADH、AAT 的氧化、还原、酯化作用下,形成青叶醛、青叶醇、己酸己酯、乙酸叶醇酯等绿叶型挥发物及带有弥散香味的果酯类[9,14]。

图1 植物中不饱和脂肪酸脂肪氧合酶代谢途径的两个支路途径Fig.1 Two branches of unsaturated fatty acid lipoxy genase pathway in plant
茶树在不同口器昆虫的危害下, 能激活LOXHPL 途径上的基因,释放GLVs 驱赶害虫,吸引天敌[15-16],与此同时,GLVs 又是茶叶加工过程中香气构成的重要组分[17]。 王晶(2010)研究了不同茶树品种加工过程中LOX 与香气形成的关系[18]。 GUI J 等(2015)以金萱为供试材料,发现脂肪氧合酶活力随着工艺过程的推进而升高,认为乌龙茶的花果香与脂肪酸氧化降解的关系更为密切[19]。 HU CJ 等(2018)以佛手和浙农 139 为试材,利用 GC-MS 与转录组测序技术, 指出LOX pathway 形成的GLVs是乌龙茶花果香品质形成过程中的主要化学组分[20]。CHO 等[21]以青心大冇为材料,研究了台式乌龙茶加工过程中关键代谢物及基因表达谱,发现在做青后期 LOX-HPL 途径的 C6 醛类、C6 醇类、C6 酸类含量均有上升。ZENG L 等以金萱为材料,探究在茶叶采后持续的机械力损伤及低温胁迫下α-亚麻酸(ALA)、亚油酸(LA)的含量及 LOX 基因相对表达量变化规律, 发现乌龙茶做青过程中的CsLOX1 基因能响应持续的机械损伤及低温胁迫[22]。 综上,GLVs 既是茶树应对外界胁迫的防御策略, 同时在讲究“看青做青”的乌龙茶加工过程中,它既是制茶过程中的重要指示物,又是乌龙茶天然花果香的前体, 但武夷岩茶加工过程中GLVs 形成及转化的研究尚未见报道。
作者采用理想化的做青工艺参数探讨了鲜叶、摇青叶、摊放叶在脂肪族类香气组分及前体物质上的差异, 认为做青过程中的机械力损伤上调了LOX、ADH 相关基因的表达量, 促使乌龙茶形成天然的花果香[9], 但仅基于 LC-MS,GC-MS 和 qRTPCR 技术分析LOX-HPL 途径上部分基因转录水平和脂肪族类香气挥发物的变化和相关性分析,缺乏了关键酶活性的探究,关键基因的选择缺乏转录组和基因组数据的支撑。 而作者以武夷肉桂实际生产中的鲜叶(F)、晒青叶(SW)、做青过程叶(1T~5T)、杀青前叶(BP)为试材,采用 GC-TOF-MS 技术,获得高通量香气挥发物, 并结合LOX-HPL 途径上关键酶LOX、ADH 及AAT 活力变化和基于转录组测序筛选获得的关键基因表达量,构建武夷岩茶加工过程中LOX-HPL 途径中“香气-酶活-基因”的代谢调控模型, 筛选LOX-HPL 途径中与香气品质形成相关的潜在基因,旨在探究实际生产加工过程中脂肪族类香气物质的形成机理[3]。
1 材料与方法
1.1 供试样品
武夷肉桂鲜叶采自武夷山慧苑坑山场,采摘时间为2018 年5 月上旬。
1.2 主要试剂
植物中脂肪氧合酶(LOX)活力检测试剂盒、乙醇脱氢酶(ADH)活力检测试剂盒及乙醇酰基转移酶(AAT)试剂盒:北京索莱宝科技有限公司产品;总RNA 抽提试剂盒: 北京天根生化科技有限公司产品; 逆转录试剂盒和实时荧光定量PCR 预混液:日本Takara 公司产品。
1.3 仪器设备
LGJ-25C 型真空冷冻干燥机:北京四环公司产品;气相色谱-质谱联用仪:德国 Gerstel 公司产品;7890B 气相色谱: 美国 Agilent 公司产品;Pegasus HT 飞行时间质谱:美国 LECO 公司产品;紫外/可见分光光度计:上海元析公司产品;高速冷冻离心机:美国贝克曼库尔特公司产品;K5500 超微量分光光度计:北京凯奥公司产品;实时荧光定量聚合酶链式反应仪:美国罗氏公司产品。
1.4 工艺过程
以武夷岩茶初制过程进行加工制作,工艺过程和主要技术参数、各工艺过程的表观叶态成像及取样节点如图1 和表1 所示。 日光萎凋(晒青)温度为20 ℃,历时30 min;采用综合做青机对萎凋叶进行做青处理,做青期间温度为26~30 ℃,做青总历时为6 h 17 min。在武夷岩茶的做青结束后,往往并未立即进行高温杀青, 而是出做青桶堆积静置1 h 左右,故增设杀青前叶。

图2 武夷肉桂加工过程中的表观叶态成像Fig.2 The apparent imaging of primary manufacturing process of Wuyi Rougui tea
1.5 分析方法
1.5.1 GC-TOF-MS 检测脂肪族类香气武夷肉桂过程样经真空冷冻干燥处理72 h 后,达“梗折即短”状态,采用研钵充分研磨呈粉末状,称取2.0 g 粉末于20 mL 顶空瓶中[7],SPME 法进行挥发性物质的提取, 通过GC-TOF-MS 对提取到的挥发性物质进行检测。

表1 武夷肉桂加工过程取样点Table 1 Sampled points during manufacturing process of Wuyi Rougui tea
SPME 条件:萃取针:PDMS/DVB(23 Ga,Plain,65 μm,美国 Supelco 公司产品);孵育温度:80 ℃;孵育时间:31 min;萃取时间:60 min;解析时间:3.5 min。
色谱条件: 色谱柱:Rxi®-5silMS (30 m×0.25 mm×0.25 μm); 进样口温度:250 ℃; 传输线温度:275 ℃;载气:氦气;氦气流量:1 mL/min;程序升温:50 ℃保持 5 min,以 3 ℃/min 的速率升至 210 ℃,保持 3 min,以 15 ℃/min 的速率升至 230 ℃;不分流进样品。
质谱条件:溶剂延迟时间:300 s;扫描范围:30~500; 检测器电压:1 530 V;EI 电离能量:70 eV;离子源温度:250 ℃。
1.5.2 紫外/可见分光光度法检测关键酶活力LOX-HPL 途径的关键酶LOX (紫外分光光度法),ADH(紫外分光光度法)及AAT(可见分光光度法)的活力检测,具体步骤均严格按照试剂盒说明书操作。
1.5.3 叶片的总RNA 提取及逆转录称取1.0 g供试材料, 利用多糖多酚植物总RNA 提取试剂盒抽提取样品的总RNA,用1.2 g/dL 变性琼脂糖凝胶电泳检测其完整性, 并对吸光度及浓度进行检测,而后将提取的总RNA 采用PrimeScript RT Reagent Kit with gDNA Eraser 逆转录为cDNA。
1.5.4 qRT-PCR 检测基因相对表达量以加工过程样的mRNA 反转录成cDNA 为模板,基于转录组测序结果,筛选出LOX-HPL 途径关键基因7 条,利用qRT-PCR 技术对关键基因的表达量进行分析。采用DNAMAN 6.0 软件设计基因的qRT-PCR 引物(表2),以 CsACT 为内标基因[23]。 反应体系、PCR 反应程序参照周子维等[9]进行。采用2-△△Ct分别计算出关键基因的相对表达量[24]。
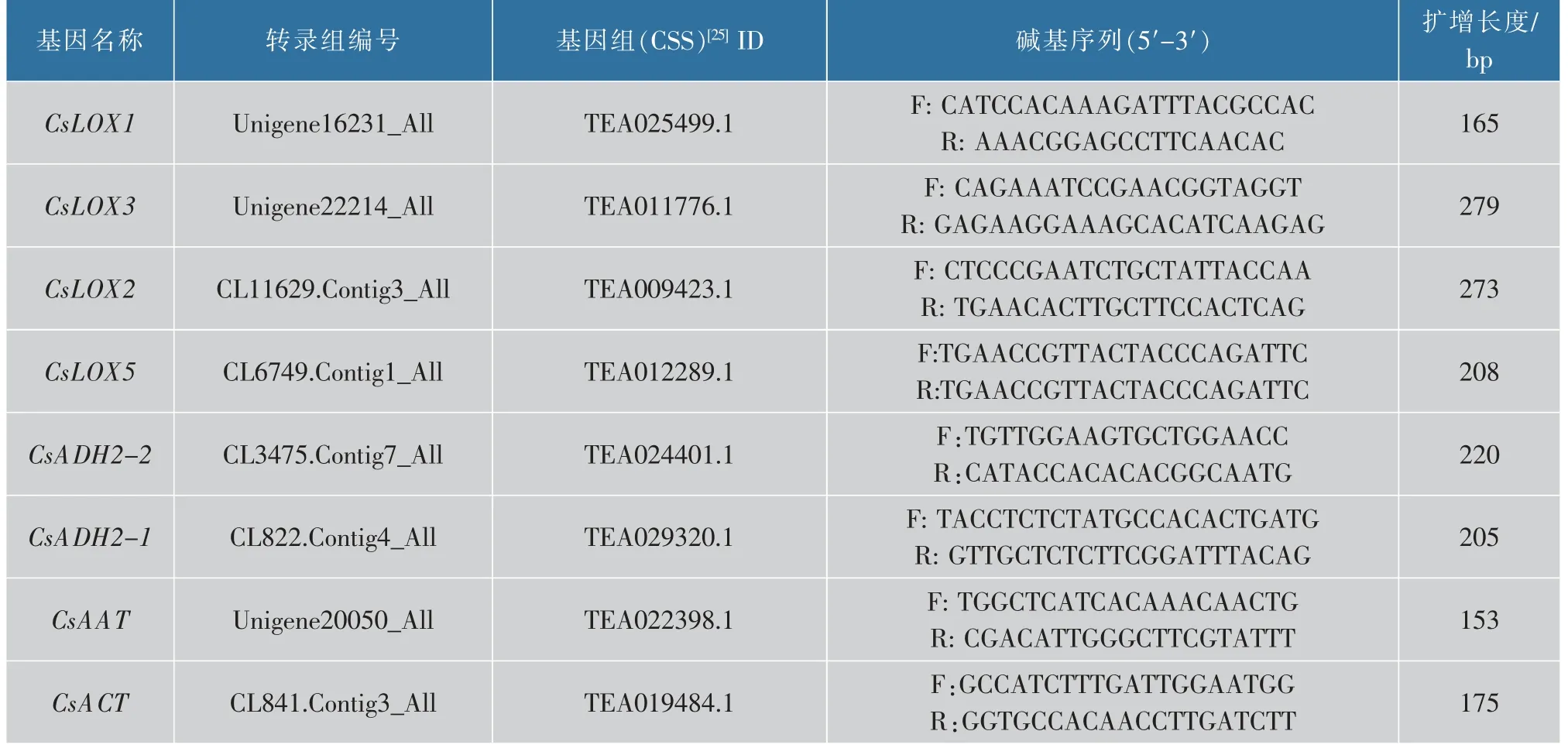
表2 茶树LOX-HPL 途径相关基因的引物列表Table 2 Primer sequences of related genes in LOX-HPL pathway in tea tree
1.5.5 数据的统计与分析差异显著性分析采用SPSS 18.0 软件中的图凯(Tukey)HSD 法,P<0.05 表示差异显著。 相关性分析采用SPSS 18.0 软件中双变量相关性检测,以皮尔森(Pearson)相关系数评估变量间的线性关系,采用双尾检验。
2 结果与分析
2.1 武夷肉桂加工中的脂肪族类香气变化
武夷肉桂加工过程中脂肪族类香气的动态变化如图3 所示,共检测获得脂肪族醛类、醇类、酸类和酯类。 脂肪族醛类为2-己烯醛,该物质质量分数在工艺过程中呈下降趋势,鲜叶在萎凋和第一次摇青后质量分数有大幅度的降低(43.20 %,59.40 %)。
脂肪族醇类有反-2-己烯醇、正己醇和反-3-己烯醇3 种, 反-2-己烯醇和正己醇的含量变化较为相似, 它们均在 5T 中累积达到最大值 (3.74×106、4.95×106)[8], 且与其他过程样品中的含量差异显著(P<0.05),而反-3-己烯醇的峰面积丰度较小,且在1T 中就达到最大值(3.96×104),在后期工艺过程中有所降低。 脂肪族酸类为己酸,它在工艺过程中呈上升趋势,在 BP 中达到最大值(3.36×106)。
脂肪族酯类较为丰富,共测得20 种,将其分为叶醇酯、己烯酯和己酸酯3 个类别,其中叶醇酯类包含苯甲酸-3-叶醇酯、丁酸甲基叶醇酯、异戊酸叶醇酯、戊酸叶醇酯、己酸叶醇酯、苯乙酸叶醇酯和柳酸叶醇酯共7 种, 除苯甲酸-3-叶醇酯在第三次摇青后再未测得外, 该类型在加工过程中变化较一致, 均是随着工艺过程的推进而逐步积累提升的,在BP 中大部分的叶醇类仍有累积,达到最大值,且与先前工艺过程中的样品间差异均达显著水平。 己烯酯类包含乙酸-2-己烯酯、乙酸-3-己烯酯、三氟乙酸4-己烯酯、顺-3-己烯基丁酯、顺-2-己烯基丁酯、顺-3-己烯酸己烯酯、反-2-己烯酸己酸酯、反-2-苯甲酸己烯酯共8 种,是脂肪族酯类中最为丰富的亚类。 在该亚类中乙酸-2-己烯酯、反-2-己烯酸己酸酯、 三氟乙酸-4-己烯酯具有类似的动态变化趋势,三者均在做青工艺中累积增多,于5T 中达最大值(1.28×106、7.04×106、5.45×105),并与先前的过程样间的差异达显著水平,值得注意的是,三者在BP 中含量出现降低,其中乙酸-2-己烯酯、反-2-己烯酸己酸酯在BP 与5T 中的差异存在显著性; 反-2-苯甲酸己烯酯、顺-3-己烯丁酯和顺-2-己烯丁酯在武夷肉桂加工过程中,具有与叶醇酯相似的动态变化规律,三者均随着工艺过程的推进不断累积增多, 在 BP 中达到最大值 (1.87×106、1.22×107、2.39×106),且与先前样品中的含量均存在显著性差异(除顺-3-己烯丁酯的BP 与5T 外)。 而剩余的乙酸-3-己烯酯在萎凋后有大幅度提升(191.10%),而随着做青工艺的开展,含量逐步下降,而顺-3-己烯酸己烯酯则呈现出先升高后降低的变化趋势,在4T中达到最大值(6.81×106),但在 5T 和 BP 中含量有所降低,但与鲜叶间的差异仍存在显著性差异。 己酸酯包含己酸甲酯、环戊基己酸酯、4-戊基己酸酯、己酸己酯、2-戊基己酸酯共5 种类型, 其中己酸甲酯、 己酸己酯、4-戊基己酸酯和2-戊基己酸酯在武夷肉桂加工过程中均呈上升趋势,除2-戊基己酸酯在 5T 中达到最大值(7.04×106)外,其余三者均在BP 中达到最大值(2.87×105、9.10×106、2.11×105),而BP 中的己酸甲酯和己酸己酯均与前期处理样品中存在显著差异, 而环戊基己酸酯在第三次摇青叶(2T)方测得(3.92×106),但 5T 和 BP 中有所降低,但与2T~4T 阶段并不存在显著性差异。

图3 武夷肉桂加工过程中脂肪族类香气物质的动态变化Fig.3 The dynamic variation of aliphatic aroma-related volatiles during manufacturing process of Wuyi Rougui tea
2.2 武夷肉桂加工过程中LOX-HPL 途径关键酶活力变化
武夷肉桂加工过程中LOX-HPL 途径关键酶LOX、ADH 及 AAT 的活性变化如图4 所示。 LOX 活力在萎凋过程中有所降低,做青阶段开始活力逐步上升并于第三次摇青叶(3T)达到最大值(1.04×104U/g),而后开始逐步下降,在5T 中达到最低值(4.7×103U/g), 且与 3T 中的活力存在显著性差异 (P<0.05), 但在做青结束后的静置摊放过程中LOX 活力又有所上升。 ADH 在工艺过程前期变化并不明显,直至 4T 中活力达到最大值(1.94 μmol/(min·g)),而后又呈下降趋势, 在 BP 中将至最低值(0.71 μmol/(min·g)), 同时与 4T 中的活力存在显著性差异(P<0.05)。 与 ADH 相似,AAT 在鲜叶(F)至 4T 中的工艺前阶段活力变化幅度较小,两两间并无显著性差异,但在5T 中,活性陡然提升至最大值(207.69 U/g), 并与其他样品中的活力均存在显著性差异(P<0.05)。
2.3 武夷肉桂加工中LOX-HPL 途径结构基因的相对表达量
采用离心柱法对武夷肉桂加工过程样进行总RNA 的抽提, 通过RNA 完整性及纯度的检测结果表明, 武夷肉桂加工过程样总RNA 的凝胶电泳均具备较清晰的18 S 和28 S 亚基条带, 且无明显的拖尾弥散,见图4(a),这表明离体的武夷肉桂鲜叶在晒青和做青处理过程中,RNA 未出现明显降解,而超微量分光光度计检测的结果表明,武夷肉桂加工过程样总RNA 的质量浓度介于400~800 ng/mL,A260nm与 A280nm比值介于 2.00~2.10 之间, 且所有样品在 A260nm处均具备单一的吸收峰,见图4(b)。 因此, 武夷肉桂加工过程样进行总RNA 符合试验要求,可开展下一步试验[10]。

武夷肉桂加工过程中LOX-HPL 途径关键基因表达谱热图如图5 所示。 LOX 相关基因CsLOX2 及CsLOX5 具有较为相似的表达模式, 均是随着工艺过程的推进先升高而后降低, 这契合了2.2 中LOX活力的变化趋势,且CsLOX2 与CsLOX5 均在2T 中达到最大值(6.96、7.53),且与其他样品的表达量存在显著性差异,CsLOX3 基因则在BP 中有所表达,CsLOX1 基因在做青前期表达略微上调后, 呈下降趋势, 同时CsLOX3 与CsLOX1 基因表达量在武夷肉桂加工过程中两两均不存在显著性差异。
CsADH2-1 基因在第四次摇青前相对表达量较低,两两间均不存在显著差异,但在5T、6T 中表达量陡然上调至最大值(2.79),且与先前处理中表达量存在显著差异。CsAAT 基因在2T、4T 中存在上调表达现象, 尽管CsLOX3 基因在BP 中表达量有所提升,但它与 CsADH2-2、CsLOX1 基因相似,在武夷肉桂加工过程中表达量变化幅度较小,两两间均不存在显著性差异。

图5 武夷肉桂加工过程中RNA 的凝胶电泳图及样品总RNA 的吸光度Fig.5 Electrophoretogram of total RNA and absorbance fluctuations of Wuyi Rougui tea samples during manufacturing process

图6 武夷肉桂加工过程中LOX-HPL 途径关键基因表达谱热图Fig.6 Heat map of LOX-HPL pathway key genes expression profile during manufacturing process of Wuyi Rougui tea
2.4 武夷肉桂加工过程中LOX-HPL 途径脂肪族类香气-酶活-基因的关联性分析
LOX-HPL 途径中脂肪族类香气、酶活力、基因表达量在武夷肉桂加工过程中的相关性热图如图7所示。
在脂肪族类香气与酶活力之间,与LOX 存在显著正相关的有顺-3-己烯酸己烯酯(0.740*)、环戊基己酸酯(0.764*),而并未发现与ADH 存在显著正相关的香气物质。
在脂肪族类香气与基因表达量之间,CsLOX1与苯甲酸-3-叶醇酯存在显著正相关 (0.800*)。 与LOX 相似,CsLOX5 基因与顺-3-己烯酸己烯酯、环戊基己酸酯也存在显著正相关(0.786*,0.740*)。 作为LOX-HPL 途径中将C6 醛还原为C6 醇的关键酶相关基因,CsADH2-1 基因与顺-2-己烯基丁酯(0.906**)、戊酸叶醇酯(0.857** )、反-2-己烯酸己酸酯(0.920**)、反-2-苯甲酸己烯酯(0.902** )、己酸己酯(0.904** )、2-戊基己酸酯(0.920**)存在极显著正相关,同时与反-2-己烯醇(0.774*)、正己醇(0.790*)、顺-3-己烯基丁酯(0.734*)、丁酸甲基叶醇酯(0.793*)、苯乙酸叶醇酯(0747* )、柳酸叶醇酯(0.825* )、 己酸甲酯 (0.734* )、4-戊基己酸酯(0.771*)存在显著正相关。 CsADH2-2 基因则与2-己烯醛存在显著正相关(0.727*),而与反-3-己烯醇存在显著性负相关(-0.796*)。
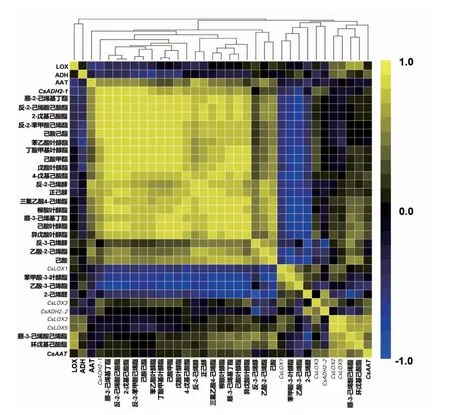
图7 武夷肉桂加工过程中LOX-HPL 途径中脂肪族类香气-酶活力-基因表达量的相关性热图Fig.7 Heat map of correlation coefficient among aliphatic aroma enzyme activity gene expression level during manufacturing process of Wuyi Rougui tea
在基因之间,CsLOX2 基因与 CsLOX5 和CsAAT 基因分别存在极显著 (0.914**) 和显著(0.762*)的正相关,同时 CsLOX5 与 CsAAT 基因亦存在显著正相关(0.732*)。
在脂肪族类香气之间,2-己烯醛与反-3-己烯醇存在极显著负相关(-0.901**)、与顺-3-己烯基丁酯(-0.761* )、己酸叶醇酯(-0.708*)、顺-3-己烯酸己烯酯(-0.785*)、柳酸叶醇酯(-0.720* )、己酸(-0.798*)间存在显著负相关;己酸与三氟乙酸4-己烯酯(0.832*)、顺-3-己烯基丁酯(0.830*)、异戊酸叶醇酯(0.712*)及己酸叶醇酯(0.822*)间存在显著正相关; 与乙酸-3-己烯酯存在显著负相关的有三氟乙酸4-己烯酯 (-0.794*)、 顺-3-己烯基丁酯(-0.724*)、顺-2-己烯基丁酯(-0.726*)、丁酸甲基叶醇酯(-0.739*)、异戊酸叶醇酯(-0.725*)、戊酸叶醇酯(-0.722*)、己酸叶醇酯(-0.777*)、反-2-己烯酸己酸酯(-0.724*)、柳酸叶醇酯(-0.757*)、己酸甲酯(-0.725*)、4-戊基己酸酯(-0.735*)、2-戊基己酸酯(-0.724*)。 而在叶醇酯、己烯酯和己酸酯的3 个亚类两两之间普遍存在着极显著或显著的正相关。 此外, 顺-3-己烯酸己烯酯和环戊基己酸酯与其他酯类间不存在显著的相关性, 但两者与LOX 及CsLOX5 存在显著正相关。
3 讨 论
3.1 武夷肉桂加工过程中的香气变化
在茶树鲜叶中,长链不饱和脂肪酸α-亚麻酸和亚油酸是LOX-HPL 代谢的前体物质[26],在LOX/HPL 酶系统的氧化裂解下,α-亚油酸降解己烯醛、己烯醇及相应的叶醇酯类,而亚油酸则降解形成正己醛、正己醇及相应的己醇酯类[27-28]。 在武夷肉桂的加工过程中,乙酸-2-己烯酯、反-2-己烯酸己酸酯、三氟乙酸-4-己烯酯等以亚油酸为前体的己烯酯类在最后一次摇青处理(5T)中达到最大值,而在杀青前的静置摊放过程中,含量有所降低。 以异戊酸叶醇酯、戊酸叶醇酯、己酸叶醇酯、苯乙酸叶醇酯和柳酸叶醇酯等7 种以α-亚油酸为前体的叶醇酯类,它们在加工过程中不断累积,在杀青前的摊放过程中亦然。 该结果说明,亚麻酸的代谢可能主要响应做青过程中所产生的外源机械力损伤,而α-亚油酸的代谢则可能不完全依赖于机械力的刺激,而主要响应鲜叶静置失水。 作者在先前的研究中,发现乌龙品种毛蟹(Camellia sinensis cv.Maoxie)在摇青叶中积累较多的己烯醇,如反-2-己烯醇(0.48%) 、反-3-己烯醇(0.30%) 以及反-4-己烯醇(3.75%),而未摇青的摊放叶中则仅有反-2-己烯醇(0.47%)[9,11]这也说表明外源机械损伤可以推进亚油酸的氧化裂解。 脂肪醛类具有强烈的青草气,在茶树鲜叶芳香油中占比约3%,作为茶叶中低级脂肪醛类的代表,2-己烯醛 (CH3CH2CH2-CH =CH-CHO, 分 子 量98.14)具备较高的沸点(150~152 ℃),是茶叶清香气味的主要成分之一[7,29]。 在武夷肉桂的加工过程中,脂肪族醛类仅测得2-己烯醛,这可能是茶树鲜叶在采摘形成的第一次机械损伤后[9]、再经回厂过程中,促进了脂肪族醛类物质转化和挥发,而仅测得较为稳定的高沸点脂肪族醛——2-己烯醛。
己醛类物质被ADH 还原为己醇的同时, 还有一部分被氧化为己酸, 而后形成己酸酯类物质,具备清新果香[30-31]。 在武夷肉桂加工过程中,测得了己酸甲酯、 己酸己酯、4-戊基己酸酯和2-戊基己酸酯等己酯类,其中己酸甲酯呈似菠萝香气,己酸己酯有生水果的清香。 己酸酯类的形成得益于己醛氧化形成的己酸。 不同于铁观音做青过程的摇青与静置,武夷肉桂的做青在综合做青机中完成,是在较为温暖(25~30 ℃)的做青间对青叶进行“送风-摇青-停机”的过程。 摇青的同时,伴随着做青设备内部风管的送风,风压将气流穿过风管的孔洞,在鲜叶加温走水的同时[32],也带来了一定的氧气,这可能是己醛类物质氧化的主要驱动力。 因此,武夷肉桂加工工艺在给离体鲜叶带来外源机械力的同时,还存在着温度、气流等的影响,许多LOX 相关基因则还可响应温度[33]、光质[34]等因素,所以基于气流、温度、 光质等因子的乌龙茶加工过程中LOX-HPL 代谢通路的研究还有待进一步深入。
3.2 武夷肉桂加工过程中的LOX-HPL 酶活力和基因转录
对多元不饱和脂肪的酶促反应,形成的C-6/-9类挥发性物质,这既是植物应对逆境胁迫的生长和生存机制[35],也是植物形成特征性风味物质的关键基础[36]。 通过对武夷肉桂加工过程中LOX-HPL 途径关键酶的活性检测, 发现随着工艺过程的推进,代谢途径的关键酶LOX、ADH 和AAT 接连在3T,4T和5T 中达到最大值,其中关键限速酶LOX 先升高后下降的结果与王晶的研究基本一致[18],而CHO 等的研究也指出东方美人茶在做青后期,脂肪族类香气的增多与LOX 活力提升有关[21]。 3T-5T 阶段作为酶活变化最为明显的时期,LOX-HPL 支路关键酶可能积极响应该时期剧烈的二次机械损伤[9]、温度[32]、失水[37]等的胁迫,而成为武夷肉桂香气形成的重要阶段。
LOX-HPL 途径相关基因在番茄[38]、桃子[39]、白梨[40]、等许多园艺植物成熟及采后处理过程中香气品质的形成起重要作用。Liu 等[17]鉴定出了茶树中首个脂肪氧合酶调控的基因CsLOX1, 该研究认为CsLOX1 基因主要在茶花的衰老过程中大量表达。近年来,基于茶树阿萨姆变种和中国变种的LOX 基因家族成员的鉴定已先后见报[32]。 在武夷肉桂的加工过程中,LOX-HPL 途径 CsLOX2、CsLOX5 基因均有上调表达,且与部分脂肪族酯类间存在极显著或显著的正相关。而作者所在课题组先前的报道[9,23]指出,在摇青处理下CsLOX1 基因表达显著上调,与青叶醛、正己醛含量呈极显著负相关,与正己醇、反-3-己烯醇、反-4-己烯醇、丁酸叶醇酯、己酸叶醇酯、苯甲酸叶醇酯、异戊酸叶醇酯及丁酸己酯含量间均存在极显著正相关, 认为CsLOX1 是做青过程中的“明星基因”,其高表达与乌龙茶的花果香有着密切相关[12]。但是,在武夷岩茶的做青过程中,CsLOX1 基因在2T 中已达到最高值, 且整个加工过程中变化并不显著。 该现象形成的原因除了茶树品种的差异外,更多的是实际生产样品与实验室处理样品的差异。 在闽北乌龙茶产区有着“勤肉桂,懒水仙”之说,这反映了武夷肉桂生产加工过程中,伴随着频繁且大量的外源机械力刺激,这在一定程度上可能推进了LOX-HPL 途径的代谢进程,进而初始CsADH2-1、CsAAT 基因在做青后期显著上调表达, 其中CsADH2-1 基因与多种具备天然花果香的酯类存在极显著或显著相关,该结果与HU C J 等[20]的研究结果较为接近,尽管CsAAT 基因与脂肪族酯类物质间未呈现出显著水平, 但该基因与CsLOX3、CsLOX5基因具有相似表达模式,它们间形成的协同共表达网络,或许正是武夷肉桂加工过程中脂肪族类香气品质形成的重要分子机理之一。
