不同牧草青贮对发酵品质的影响
2020-12-28滕道明郝明静
滕道明,郝明静
(江苏省铜山中等专业学校,江苏徐州 221116)
青贮发酵取决于不同微生物群之间的竞争,所需要的微生物中乳酸菌通常在青贮过程中起主导作用,而一些不需要的微生物也会生长,造成厌氧或好氧腐败。肠球菌、乳球菌和片球菌通常被认为能启动青贮发酵,而在后期,更耐酸的乳酸菌如植物乳杆菌和短乳杆菌可能取代它们(张庆等,2014)。测定发酵产物的组成可以大致评价发酵的成功程度。如果青贮饲料中乳酸含量较高,则可有效抑制非乳酸菌种类微生物的增殖,减少贮藏过程中干物质的损失和蛋白质的降解(吴寒,2015)。相关微生物的信息有助于了解如何优化青贮过程以提高生产率,因为除了乳酸菌外,各种微生物还可以生产主要的发酵产物,如乳酸、乙酸和乙醇。为了提高发酵可接受性,最好避免高水分的原料,但在大规模青贮中,高水分作物可以用来节省时间。此外,应分析不同环境的牧草和谷物青贮饲料,以了解不同作物品种之间青贮菌群落的差异,从而解释发酵模式的差异(Nussion,2005)。因此,本研究旨在评估3 种牧草在不同青贮发酵和需氧变质时期下对其青贮品质的影响。
1 材料与方法
1.1 牧草青贮将收割的黑麦草、羊草和全珠玉米在不添加任何添加剂的条件下直接切段约12 mm长,在实验仓中青贮发酵3、7、14、28、56 和112 d(一式三份)。青贮饲料在第14、28、56 和112 天完全打开后,一半含量(150 g 左右)放入一个500 mL 聚乙烯瓶。瓶子的顶部没有盖住,在25℃的房间里暴露7 d。
1.2 化学分析分别在开仓时和好氧稳定性试验后采集青贮饲料样品。在60℃的烘箱中干燥48 h 来测定干物质含量。青贮饲料的pH、乳酸、短链脂肪酸和醇的含量参考Li 和Nishino(2011)的方法进行。用800 g/L 乙醇提取水溶性碳水化合物,用苯酚-硫酸法测定其含量。
1.3 统计分析对数据采用SAS 软件单因素模型进行方差分析,采用Tukey 法进行多重比较。采用T 检验测定青贮仓开仓时青贮仓内微生物种群与经好氧稳定性试验后青贮仓内微生物种群的差异。P <0.05 表示差异显著。
2 结果与分析
2.1 牧草青贮前化学成分由表1 可知,黑麦草、羊草和全珠玉米的干物质含量分别是210、168和251 g/kg,水溶性碳水化合物含量分别是125、18.5 和70.5 g/kg。青贮前羊草和全珠玉米附带的乳酸菌数量大于106CFU/g,而黑麦草乳酸菌数量低于检测限值(102CFU/g)。
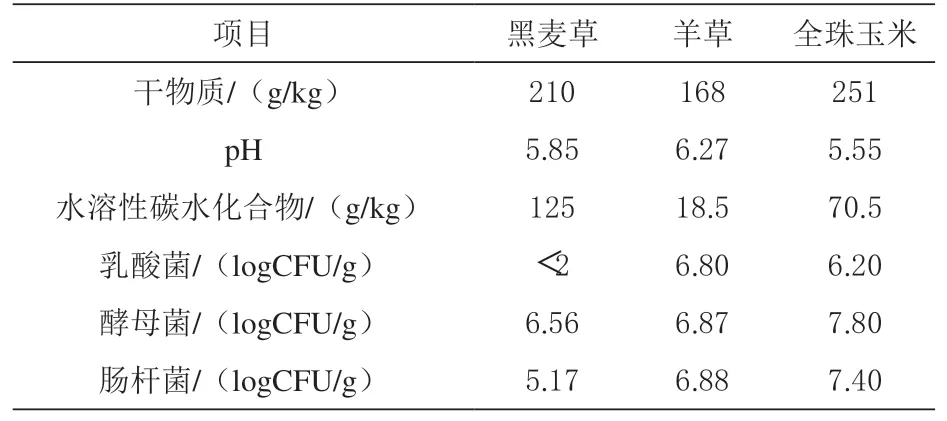
表1 几种牧草青贮前化学成分
2.2 青贮发酵和耗氧变质对牧草青贮品质的影响由表2 可知,丁二醇和乙醇是黑麦草青贮的主要发酵产物。青贮3 d 时,丁二醇和乙醇的含量分别为58.2 和27.7 g/kg,3 ~7 d 时再增加约10 g/kg,而乙醇含量变化不大。在青贮过程中,乳酸含量从青贮第3 天的低水平升高到28 d 的最大值(P <0.05)。丁酸从青贮开始就一直在生成,在青贮122 d 时,丁酸产量大于乳酸。无论青贮期如何,乳酸菌计数均在107cfu/g。虽然在青贮28 d 前酵母菌计数大于107cfu/g,但在第56 天后,其计数低于可检测水平。同样,肠杆菌的水平在第14 天大于107cfu/g,之后在第28 天下降到105cfu/g,在第56 天计数低于可检测水平。
由表3 可知,在羊草青贮饲料中乙酸是主要产物。其中青贮3 d 时为26.0 g/kg,青贮112 d 时为57.8 g/kg。青贮初期乳酸含量仅次于乙酸,但在第14 天显著下降(P <0.05)。丁二醇的含量在青贮过程中没有显著变化(P >0.05)。另一方面,随着青贮时间的延长,乙醇含量显著增加(P <0.05),同时丁酸的含量也相应提高,直到青贮14 d 才检测到,但从28 d 开始产量有了大幅提高。无论青贮期如何,均未检测到乳酸菌、酵母菌和肠杆菌。
由表4 可知,全珠玉米青贮饲料发酵初期乳酸产量显著提高(P <0.05),其中青贮3 d 其乳酸含量为50.4 g/kg,在青贮28 d 时达到峰值。虽然乙酸含量保持在乳酸含量的1/3 以下,但在青贮56 d 后 显 著 增 加(P <0.05),在112 d 时 达到最大值。从发酵开始到发酵结束,乙醇含量基本稳定(P >0.05)。青贮3 d 时乳酸菌计数最 高(P <0.05),青 贮112 d 时 降 至106cfu/g。同样,随着青贮期的延长,酵母菌数量显著减少(P <0.05)。
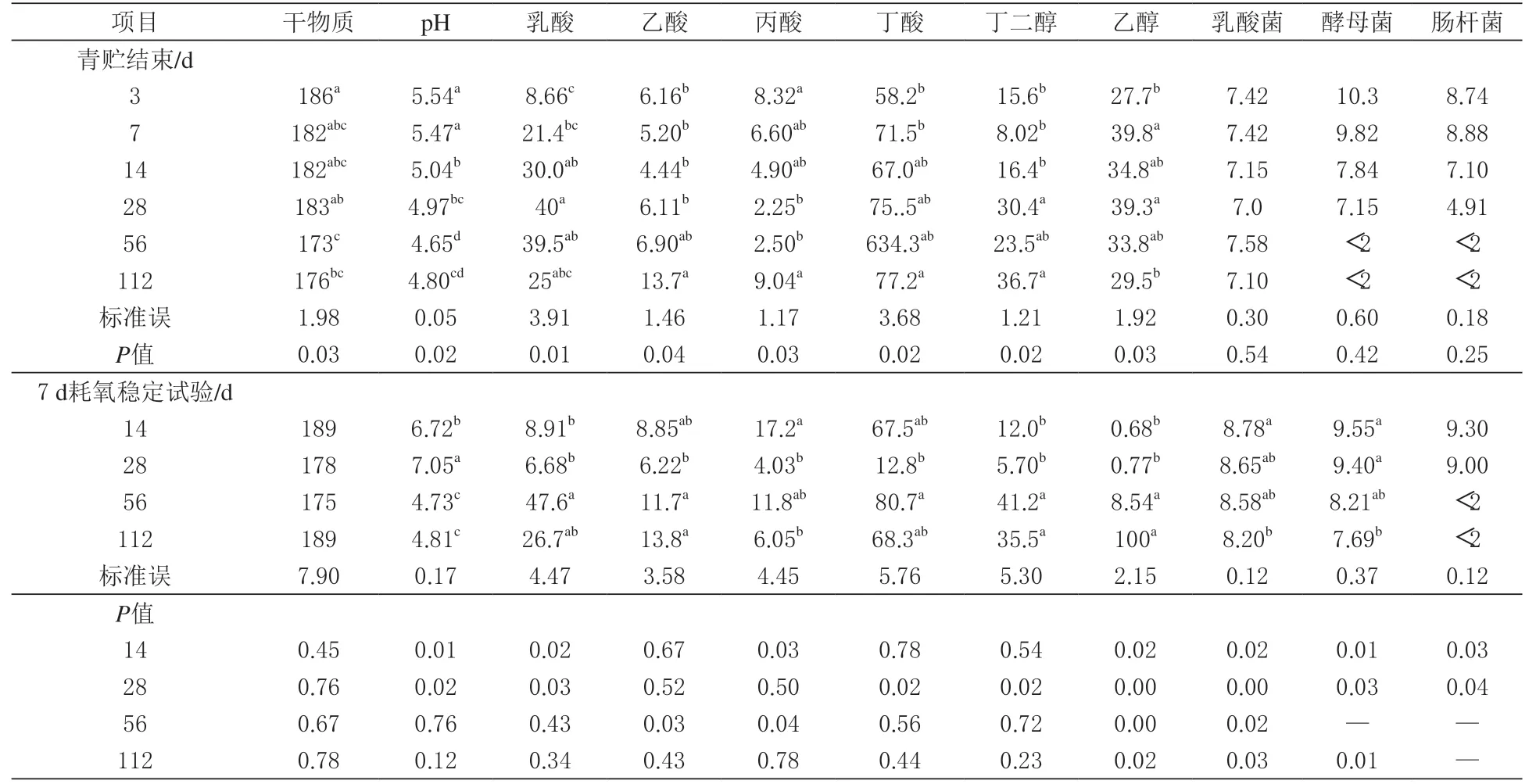
表2 青贮发酵和耗氧变质对黑麦草青贮品质的影响
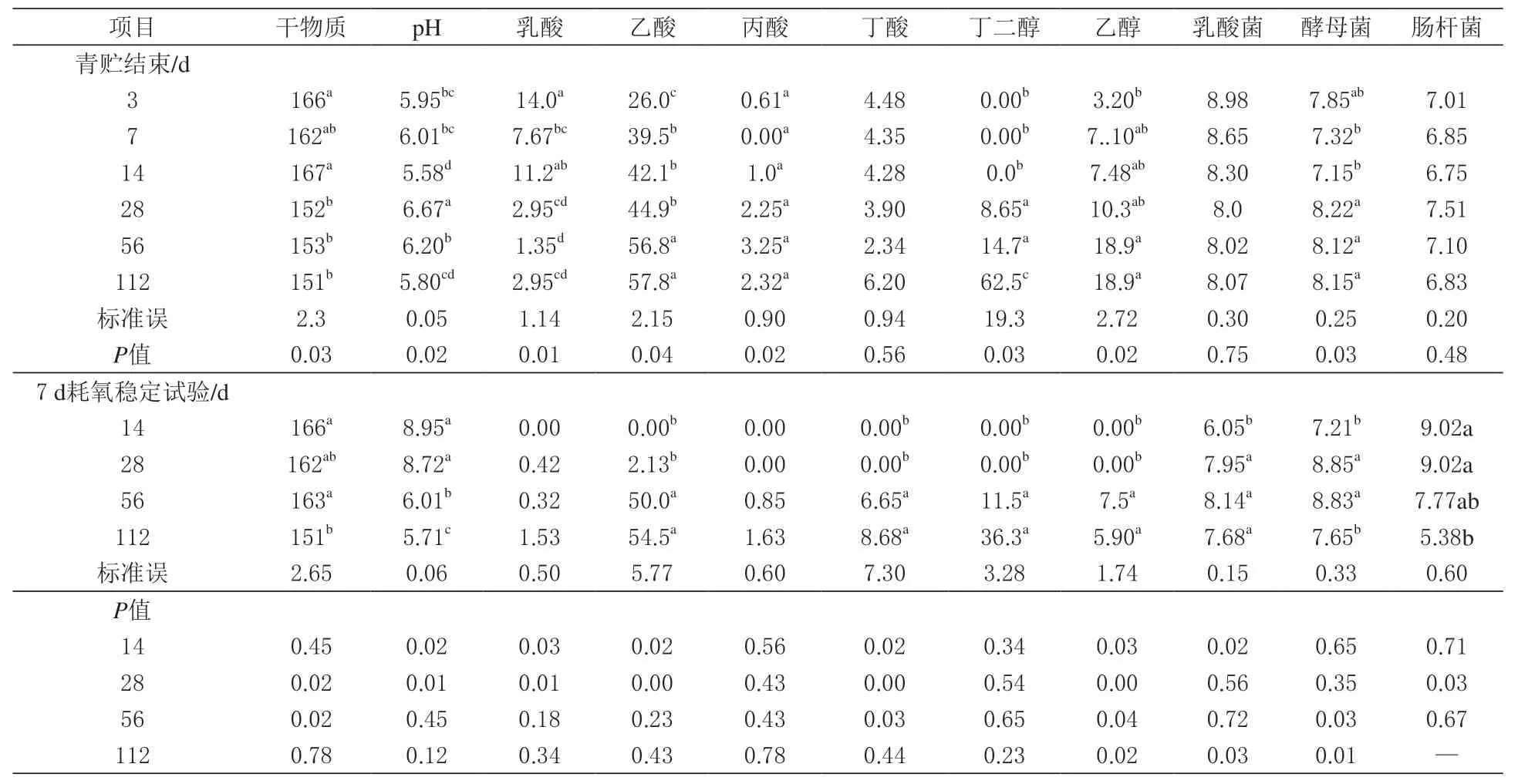
表3 青贮发酵和耗氧变质对羊草青贮品质的影响
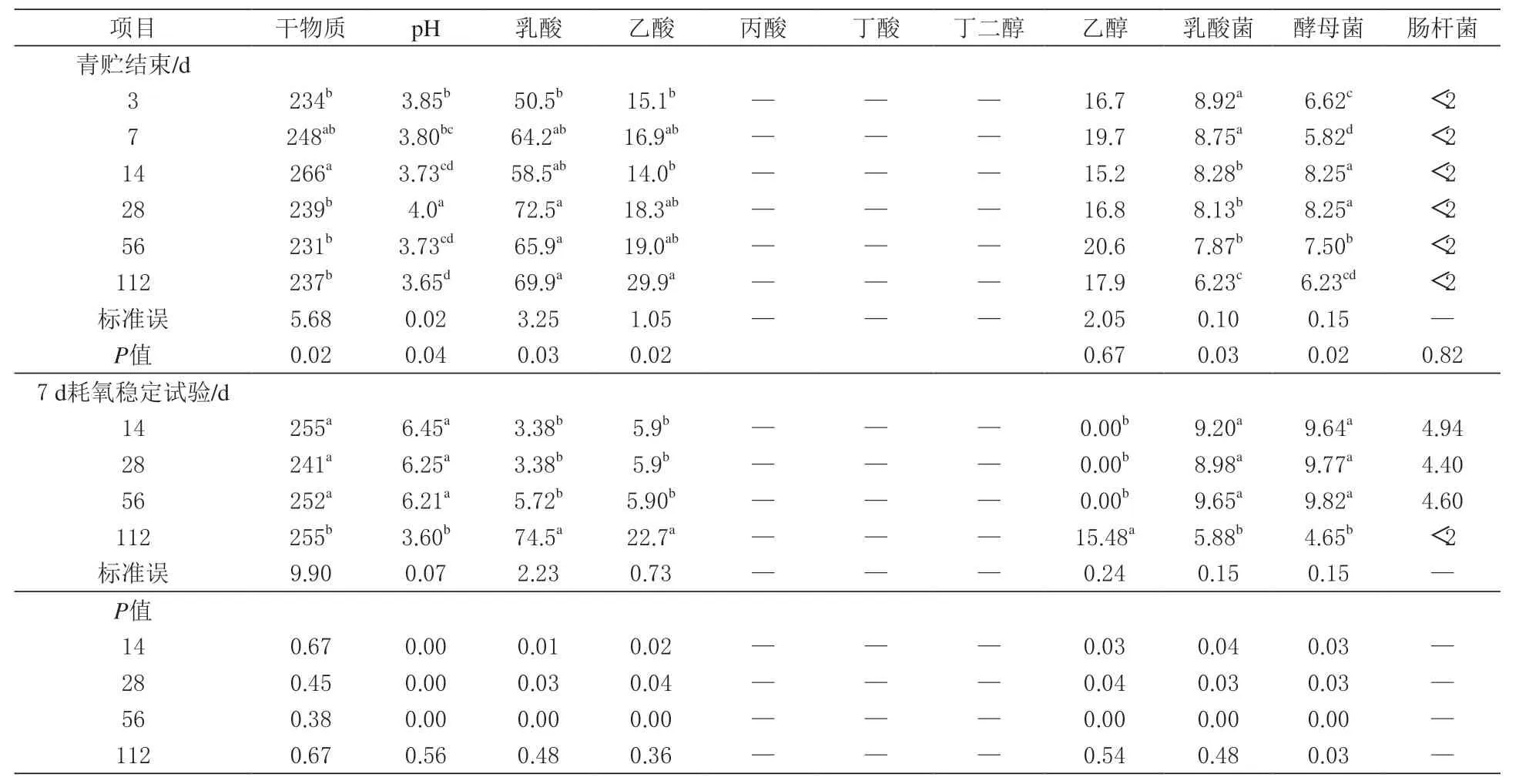
表4 青贮发酵和耗氧变质对全珠玉米青贮品质的影响
3 讨论
虽然在青贮前的作物中发现了足够的水溶性碳水化合物,但黑麦草青贮中丁二醇和乙醇的产量得到提高,而在全珠玉米青贮中有较高水平的乳酸,同时试验检测的细菌群落数据与这些差异一致。在黑麦草青贮饲料中,从发酵开始就检测到肠杆菌。值得注意的是,青贮前黑麦草中发现的肠杆菌很快被适应青贮环境的肠杆菌所取代,意味着虽然在最初青贮的3 d 增菌过程中肠细菌数量的增加很小,但群落结构发生了很大变化。
Parvin 和Nishino(2009)报道,在热带牧草青贮中,主要的发酵产物往往是乙酸而不是乳酸。高水分制备和长时间的青贮可能会加剧乙酸发酵(Nishino 等,2012)。但从发酵一开始,与提高乙酸产量相关的细菌并未检测到。青贮3 和7 d,乳酸和乙酸含量发生显著变化,这可能与植物乳杆菌参与延长青贮导致的乙酸含量增加有关(Nishino,2005),因为植物乳杆菌可以在缺糖条件下将乳酸代谢为乙酸。同样的代谢过程在本研究中也可能发生,因为在青贮初期检测到了植物乳杆菌。乙酸含量的增加远远大于乳酸含量的减少,因此,乳酸代谢可能是促进乙酸发酵的原因。
青贮饲料中的乙酸、丙酸和丁酸会抑制酵母菌和霉菌,因此,黑麦草和羊草青贮饲料中乙酸和丁酸的增加和全珠玉米青贮饲料中乙酸的增加可以作为抑制微生物的物质。由于羊草青贮3 d 后显示出较高的乙酸含量(>26 g/kg),因此,并不是14 和28 d 青贮饲料被破坏的原因。相比之下,全珠玉米青贮112 d 后pH 为3.65,因此,未解离的乙酸为>0.90。对于微生物的抑制非解离酸最重要,而解离分子的作用是次要的(Muck 等,1991)。因此,本研究的结果再次证实了非解离酸在抑制腐败中的重要性。
4 结论
丁二醇和乙醇是黑麦草青贮的主要发酵产物,乙酸是羊草青贮的主要产物,全珠玉米青贮饲料发酵初期乳酸产量显著提高,其中青贮3 d 其乳酸含量为50.4 g/kg,在青贮28 d 时达到峰值。
