日本血吸虫正常发育和发育阻遏雌虫的转录组差异分析
2019-11-11程贵凤李肖纯秦芳林何川川刘金明古少鹏金亚美
程贵凤,李肖纯,秦芳林,何川川,刘金明,古少鹏,金亚美
(1.山西农业大学动物科技学院,山西太谷030801 ; 2.中国农业科学院上海兽医研究所 农业部动物寄生虫学重点开放实验室,上海闵行200241 ; 3.上海师范大学生命与环境科学学院,上海徐汇200233)
血吸虫病是由血吸虫感染引起的一种人兽共患寄生虫病,感染者超过2亿,有将近8亿人面临着感染风险[1-3],在我国主要流行的是日本血吸虫病。血吸虫是雌雄异体[4],雌虫只有与雄虫持续不断地合抱才能促进和维持卵黄腺和卵巢的发育与成熟,才能正常产卵[5]。成熟雌虫所产大量虫卵随宿主粪便排出体外造成血吸虫病的流行,同时沉积在宿主体内的虫卵会引起严重的病理损害[6]。而单性感染的雌虫,缺乏与雄虫的接触,生殖系统发育停滞,不能正常产卵。此外,当配对的雌虫与雄虫分离,雌虫将退回到未成熟状态并停止产卵[7-8]。因此,研究与血吸虫生殖发育和产卵相关的通路和基因,深入了解日本血吸虫雌虫性成熟和产卵的调控机制,对控制该病的发生和传播十分重要。
本研究用RNA-seq高通量测序技术对日本血吸虫25日龄MF和SF的转录组进行测序、定量及差异表达基因的筛选,利用生物信息学分析差异表达基因,寻找与雌虫生殖发育和产卵相关的基因,以期为研究日本血吸虫雌虫的生长发育、性成熟和产卵机制提供理论依据。
1 材料与方法
1.1 材料
1.1.1 主要试剂与仪器 Prime-ScriptRTreagent Kit with gDNA Eraser、SYBR®Premix ExTaqTMⅡ,购自TaKaRa生物工程有限公司;TRIZol®试剂,购自上海英潍捷基生物公司;ABI 7500实时荧光定量PCR仪(美国ABI公司)。
1.1.2 实验动物与虫株 日本血吸虫中国大陆株尾蚴由中国农业科学院上海兽医研究所血吸虫病研究室提供;4~6周龄雄性BALB/c小鼠,购自上海斯莱克实验动物有限责任公司。
1.2 方法
1.2.1 25日龄MF和SF的收集 采用腹部贴片法感染小鼠,于感染25 d剖杀,以肝门静脉灌注法收集合抱分开的雌虫;以单钉螺逸出的尾蚴感染小鼠,25 d剖杀,结合形态观察,收集SF虫体,SF较MF小,所收集虫体用PBS洗涤3 次,液氮罐冻存备用。
1.2.2 RNA的提取及日本血吸虫mRNA文库的构建 按照TRIZol说明书提取总RNA,并用NanoDrop1000分光光度计评估RNA的质量。总RNA样品用DNA酶I处理以降解任何可能的DNA污染,使用Oligo(dT)磁珠来富集mRNA,向得到的mRNA中加适量片段化缓冲液,在高温条件下使其片段化,再以片段化后的mRNA为模板, 合成cDNA,经磁珠纯化、末端修复、3′末端加碱基A、加测序接头后,进行PCR扩增,从而完成整个文库制备工作,并进行测序。
1.2.3 参考基因组比对及测序评估 将测序结果与日本血吸虫基因组数据库比对并进行测序饱和度分析,通过对比对数据的统计和分析,判定该数据是否可用于后续分析。
1.2.4 差异表达基因筛选 基因表达量的计算采用RPKM方法消除基因长度和测序量差异对计算基因表达的影响,得到的基因表达量直接用于比较不同样品间的基因表达差异,公式为:RPKM=106C/(NL/103),RPKM为基因表达量,C为唯一比对到该基因的读取数,N为唯一比对到参考基因的总读取数,L为该基因编码区的碱基数。以│Log2Ratio│≥1, p-Value<0.001为标准筛选差异表达基因,其中错误检出率小于0.001。
1.2.5 生物信息学分析 用BLAST将检测到的基因与Nr(Non-redundant protein sequence database in GenBank)数据库进行比对,获得注释信息。利用Blast2GO软件将基因序列与GO(Gene ontology)数据库进行比对,从生物学进程,细胞组分和分子功能这3个层面进行基因GO注释,并筛选出差异基因富集的显著性GO条目;同时将基因于KEGG(Kyoto encyclopedia of genes and genomes)数据库进行通路注释,获得差异表达基因相应的通路信息。
1.2.6 荧光实时定量RT-PCR(qRT-PCR)检测差异基因的转录水平 为验证测序结果的可靠性,从MF和SF差异表达的基因中随机选取5个上调基因和5个下调基因进行qRT-PCR检测。用Primer 5.0设计这些基因的特异性引物,见表1(由上海英潍捷基生物公司合成)。提取25 d MF和SF虫体的总RNA,按Prime-ScriptRTreagent说明书反转录为cDNA,并按SYBR®Premix ExTaqTMⅡ说明书进行qRT-PCR,每个反应均做三孔重复。利用ABI 7500 Software v2.0.5分析结果,以tublin基因作为内参,得出相对于每百万持家基因的目的基因含量。
1.2.7 统计学分析 用GraphPad Prism 5进行数据分析,差异表达基因在MF与SF的差异性分析用t-test进行检验;P-values≤0.05或0.01视为差异显著或极显著。

表1 qRT-PCR检测的基因序列和扩增长度
2 结果与分析
2.1 原始数据分析 从MF和SF中分别获得3 696 376和3 662 109条clean reads,且clean reads占测序总读数的99.5%,样品质量良好。将clean reads数匹配到日本血吸虫基因组数据库,匹配率达79%左右。
2.2 测序饱和度 测序饱和度结果显示,当测序量达到一定值时,25天MF和SF中鉴定到的基因数量不再增加,说明测序结果是全面且饱和的,可进行后续生物学分析。
2.3 差异表达基因的筛选 RNA-seq共检测到11 171个基因。在这些基因中,以│Log2Ratio│≥1,p-Value<0.001为条件,筛选到775个在MF和SF之间显着差异表达的基因,其中401个基因在MF中上调, 374个基因在SF中上调。
2.4 GO功能分析 从BP(Biological process, 生物学进程)、CC(Cellular components, 细胞组分)、MF(Molecular function, 分子功能) 3个层面进行GO注释,如图1所示,在生物学进程中,差异表达基因更多的参与细胞进程,代谢进程和单一生物进程。在细胞组分中,差异表达基因集中在细胞、细胞器和细胞膜中。在分子功能中,大多数差异表达基因与结合和催化活性相关。
GO功能富集分析还结合了表达模式的聚类分析,如表2所示,差异表达基因的主要生物学功能集中在:氧化磷酸化,离子跨膜转运,血红素结合和铁离子结合。

图1 差异基因的GO注释
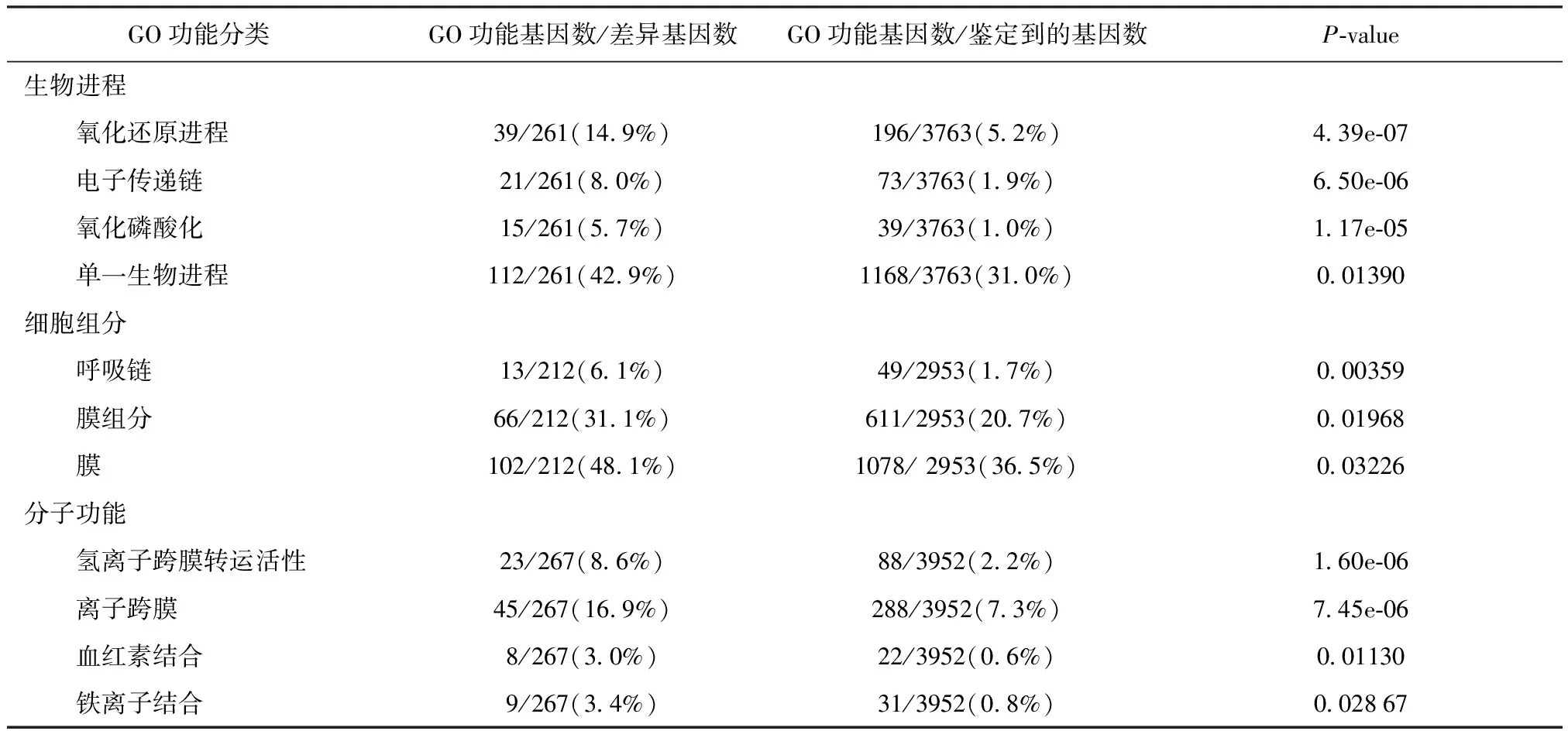
GO功能分类GO功能基因数/差异基因数GO功能基因数/鉴定到的基因数P-value生物进程 氧化还原进程39/261(14.9%)196/3763(5.2%)4.39e-07 电子传递链21/261(8.0%)73/3763(1.9%)6.50e-06 氧化磷酸化15/261(5.7%)39/3763(1.0%)1.17e-05 单一生物进程112/261(42.9%)1168/3763(31.0%)0.01390细胞组分 呼吸链13/212(6.1%)49/2953(1.7%)0.00359 膜组分66/212(31.1%)611/2953(20.7%)0.01968 膜102/212(48.1%)1078/ 2953(36.5%)0.03226分子功能 氢离子跨膜转运活性23/267(8.6%)88/3952(2.2%)1.60e-06 离子跨膜45/267(16.9%)288/3952(7.3%)7.45e-06 血红素结合8/267(3.0%)22/3952(0.6%)0.01130 铁离子结合9/267(3.4%)31/3952(0.8%)0.02867
2.5 差异表达基因的KEGG分析 本研究将所有显著差异基因与KEGG数据库进行比较,发现这些差异基因富集到206个途径。其中显著性富集途径涉及到代谢途径,肌动蛋白细胞骨架的调节,溶酶体等,见图2。
2.6 差异表达基因的验证 为了验证RNA-seq结果,从鉴定到的差异基因中随机选择5个上调基因和5个下调基因用于qRT-PCR验证,结果显示,这些基因在转录水平上也表现出相应的上调或下调,见图3A和3B。
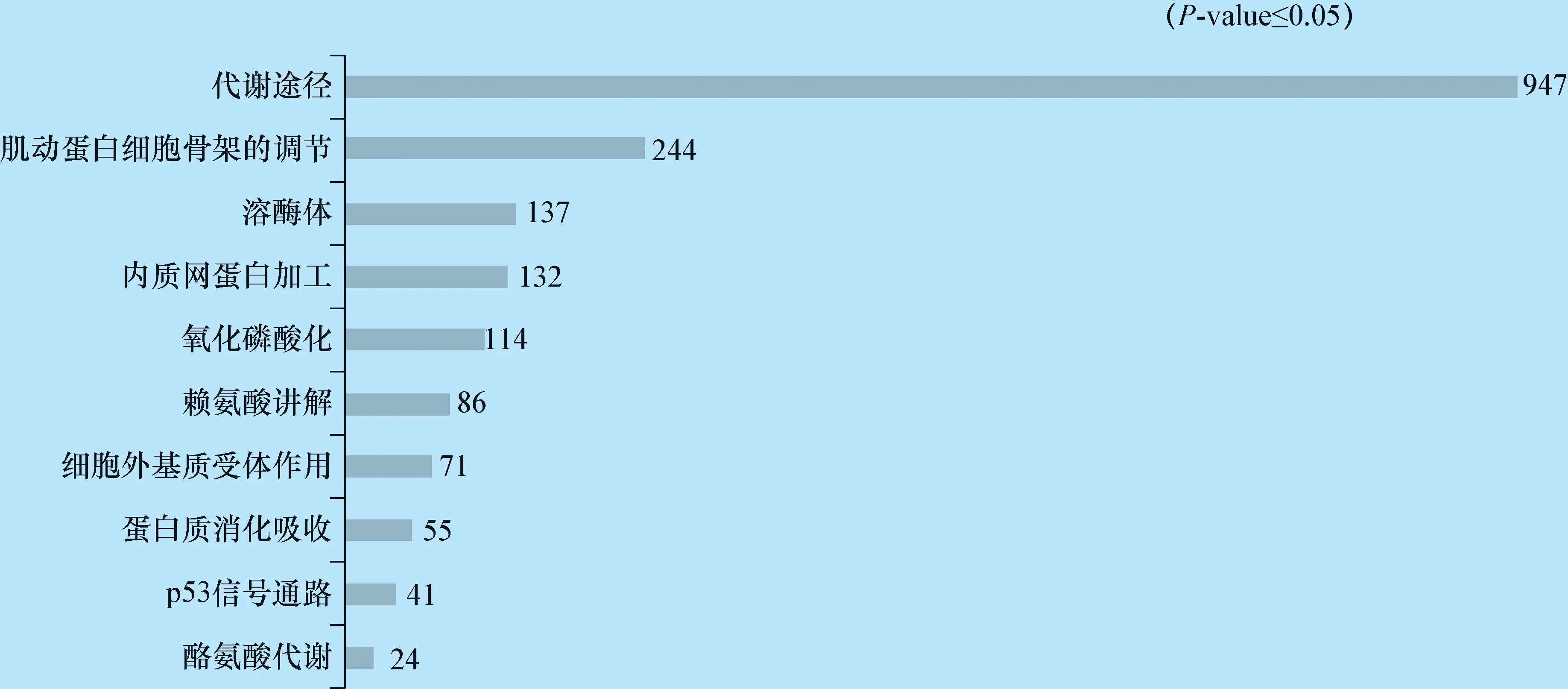
图2 差异表达基因的KEGG分析
3 讨论
为探索血吸虫雌虫的生长发育、产卵等机制,本研究使用RNA-seq技术对25天MF和SF的样品进行了大规模测序、定量及差异表达基因的筛选。经测序饱和度分析表明测序结果是饱和且全面的,全部的clean reads数与日本血吸虫基因组序列匹配发现两组样本的匹配率皆达79%以上,但仍有20%未知标签,这表明存在新的基因需进一步鉴定。
日本血吸虫雌虫的性成熟和产卵对血吸虫病的发生和传播至关重要,为鉴定与这些生物学过程相关的基因,本研究测序后以│Log2Ratio│≥1,p-Value<0.001为条件,筛选出775个显着差异表达的基因,其中MF中有401个上调基因,SF中有374个基因上调。生物信息学分析表明,鉴定的差异基因主要参与代谢过程,生物合成,细胞和运动过程。
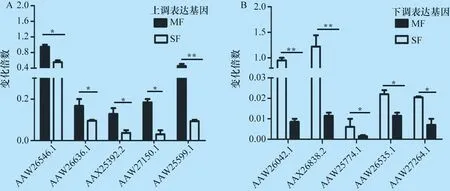
图3 qRT-PCR验证差异表达基因结果
*:P≤0.05 ; **:P≤0.01
代谢过程对血吸虫的生长发育十分重要,本研究KEGG富集分析发现,两组样本中差异表达基因富集到的最多参与途径是代谢过程,且参与这些代谢过程的差异基因更多地在MF中上调表达,涉及的代谢过程包括核苷酸代谢,氨基酸代谢和葡萄糖代谢。本研究富集分析发现参与编码腺苷磷酸化酶、鸟苷酸激酶、天门冬氨酸氨基甲酰转移酶和二氢转移酶的基因在MF中上调,且已证明这些酶在日本血吸虫核苷酸代谢的自我合成途径中起重要作用[9-11],这说明核苷酸代谢可为日本血吸虫雌虫提供所需的能量。同时,检测到参与编码酪氨酸酶和精氨酸酶的基因在MF中高表达,已有研究表明,酪氨酸可促进雌虫产卵[12],而精氨酸酶有助于雌虫自身的胶原沉积[13]。此外,葡萄糖代谢对日本血吸虫的能量需求非常重要,它是血吸虫生长和发育的主要来源[14]。本研究检测到很多参与糖酵解的酶,以及参与编码胰岛素蛋白的基因在MF中上调,这表明雌虫的生长发育与葡萄糖代谢提供的能量是不可分割的。
蛋白质的合成在维持日本血吸虫的正常运动和满足雌虫产卵方面发挥重要作用,本研究检测到差异基因编码的大量蛋白在MF中上调,如膜相关蛋白,它们可调节囊泡的融合和对接[15],并在血吸虫葡萄糖吸收方面起重要作用[16]。还有热休克蛋白,它们能保护血吸虫免受有害环境的影响[17]。此外,差异表达基因GO富集分析发现,血红素结合和铁结合过程在MF中显着上调,研究表明,血吸虫可以利用自身的组织蛋白酶降解宿主的血红蛋白为珠蛋白和血红素,提供所需的营养来源,且成熟雌虫的卵黄腺中储存有大量的铁,可促进雌虫的产卵[18]。
KEGG富集分析和GO功能注释发现在SF中上调表达的基因多参与细胞通讯和运动,如参与编码肌动蛋白和肌球蛋白的基因,这些基因的高表达说明发育阻遏雌虫运动系统发达,为与雄虫合抱做准备。
4 结论
本研究分析了MF和SF之间的转录组差异,发现许多与雌虫生长发育和产卵有关的差异基因和通路,为研究日本血吸虫雌虫的生殖发育机制提供基础。
