二氢杨梅素对Bloom解旋酶结构和生物学活性的影响
2019-09-24刘金河张望明晏文涛许键炜葛章文
刘金河,张望明,晏文涛, 许键炜,葛章文
(1. 贵州医科大学细胞工程生物医药技术国家地方联合工程实验室,组织工程与干细胞实验中心,贵州省再生医学重点实验室,贵州 贵阳 550004; 2. 中国医学科学院成体干细胞转化研究重点实验室;3. 贵州省人民医院检验科,贵州 贵阳 550002;4. 贵州中医药大学第一附属医院检验科,贵州 贵阳 550001)
Bloom综合征(Bloom syndrome,BS)是一种隐性的常染色体遗传病,临床主要特征是生长发育迟缓、面部有对光敏感的斑、免疫缺陷、易患多种癌症及不孕不育[1]。BS主要是由细胞内的Bloom解旋酶(Bloom helicase,BLM)突变导致基因组的不稳定,进而导致各种基因突变发生并诱发癌症。BLM解旋酶能够结合双链DNA,利用ATP提供能量,由3′-5′解开双链DNA[2]。因此,研究小分子药物对BLM解旋酶功能的影响,寻找靶向特效抑制药物,对治疗此类癌症具有重要意义。本研究中使用的BLM解旋酶为该酶的核心区域,氨基酸残基序列区间为642-1290,简称BLM642-1290。
二氢杨梅素(dihydromyricetin,DMY)是一种黄酮类化合物(结构见Fig 1),主要存在于杨梅科、柳科、葡萄科、藤黄科等植物中,在藤茶属植物中含量最高[3-4]。最近的研究表明,DMY具有广泛的药理活性,能够通过促进细胞凋亡,多环节阻滞或延缓肿瘤的发生发展[5-6]。在急性胰腺炎(acute pancreatitis,AP)的损伤修复中,通过抑制TRAF3-p38信号通路来抑制AP的应答[7]。在对乙酰氨基酚(acetaminophen,APAP)诱导的小鼠急性肝损伤中,具有良好的修复作用[8]。在抑制肝脏的脂肪堆积,防止动脉粥样硬化方面,也具有明显的疗效[9]。在临床上,能缓解地塞米松造成的肌肉萎缩[10], 能通过抑制semaphorin 4D来抑制结直肠癌的发展[11]。另外,DMY的抗氧化作用也比较强,在抗炎、抗病原微生物、解酒护肝、调血脂等方面也有广泛的应用,具有较大的临床应用潜力[12]。
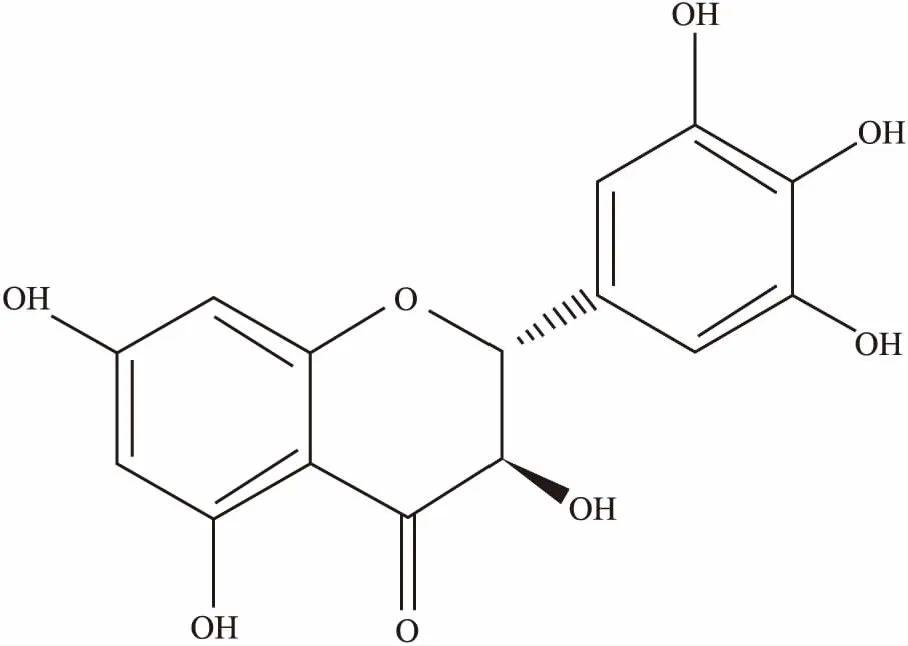
Fig 1 Chemical structure of DMY
目前对DMY的研究有很多,主要关注DMY的各种药理作用,其中抗肿瘤方面着重在肝癌、乳腺癌、卵巢癌等恶性肿瘤方面,而DMY对这些癌细胞及活体影响的分子机制的研究却鲜见报道。本文采用圆二色谱(circular dichroism,CD)、自由磷检测、紫外光谱、荧光偏振等技术和方法,研究DMY对BLM642-1290解旋酶的二级结构、生化活性、构象的影响,探讨DMY与BLM解旋酶相互作用的分子机制。
1 材料与方法
1.1 试剂DMY(CAS27200-12-0,20 mg,上海源叶生物公司);ATP酶活性检测试剂盒(英国Innova Biosciences公司);DNA底物杂交缓冲液(20 mmol·L-1Tris-HCl,100 mmol·L-1NaCl,pH 7.9);荧光偏振实验反应缓冲液(20 mmol·L-1Tris-HCl,25 mmol·L-1NaCl,3 mmol·L-1MgCl2和0.1 mmol·L-1DTT,pH 7.9);ATP酶活性检测缓冲液(0.1 mol·L-1Tris-HCl,5 mmol·L-1MgCl2,1 mmol·L-1ATP和4 nmol·L-1ssDNA,pH 7.9);紫外光谱实验缓冲液C(20 mmol·L-1Tris,500 mmol·L-1NaCl,500 mmol·L-1咪唑,10%甘油,pH 7.9);圆二色谱实验缓冲液为1×PBS。其他化学试剂均为国产分析纯。
1.2 仪器Ultrospec2100 Pro紫外分光光度计(美国GE公司);NGCTM蛋白质纯化系统(美国Bio-Rad公司);Beacon 2000荧光偏振仪(美国Panvera公司);Milli-Q 超纯水系统(美国Millipore Corp);Bio-Logic电化学工作站(法国,Bio-Logic)。
1.3 DNA底物实验中的DNA底物由北京鼎国工程技术有限公司合成,其长度和序列见Tab 1。在40 μL DNA杂交缓冲液中,分别加入5 μL 100 μmol·L-1A1和A2单链DNA(ssDNA, A2),90 ℃水浴3 min,室温下自然冷却复性,获得双链DNA(dsDNA,A1A2),其作为荧光偏振实验中检测所需的双链DNA底物。ATP酶活性测定中使用A1作为DNA底物。
1.4 BLM642-1290解旋酶的制备BLM解旋酶的重组大肠杆菌由西北农林科技大学奚绪光教授馈赠。首先,将BLM642-1290解旋酶大肠杆菌的菌种在37 ℃复苏,然后接种到5 L培养基中进行扩大培养;在18 ℃条件下,扩大培养后的菌种中加入终浓度0.4 mmol·L-1IPTG诱导表达20 h; 4 ℃条件下,4 000 r·min-1离心15 min,收集管底部菌体,加入适量buffer A(20 mmol·L-1Tris,500 mmol·L-1NaCl,0.5 mmol·L-1咪唑, 10%甘油,pH 7.9),高压细胞破碎仪(英国Constant Systems公司)进行细胞破碎(压力为21 KPSI),破碎1次,离心收集上清液; 在蛋白纯化仪上,用镍亲合层析柱和Superdex-200(美国GE Healthcare公司)进行分离和纯化,获得可用于开展生物学活性研究的BLM642-1290解旋酶(Fig 2),考马斯亮蓝染色检测BLM642-1290蛋白纯度(Fig 3)。本实验所需蛋白为Fig 3中1号泳道BLM,BCA法测定BLM642-1290解旋酶蛋白浓度为0.383 g·L-1。

Tab 1 Oligonucleotide sequences used in this study
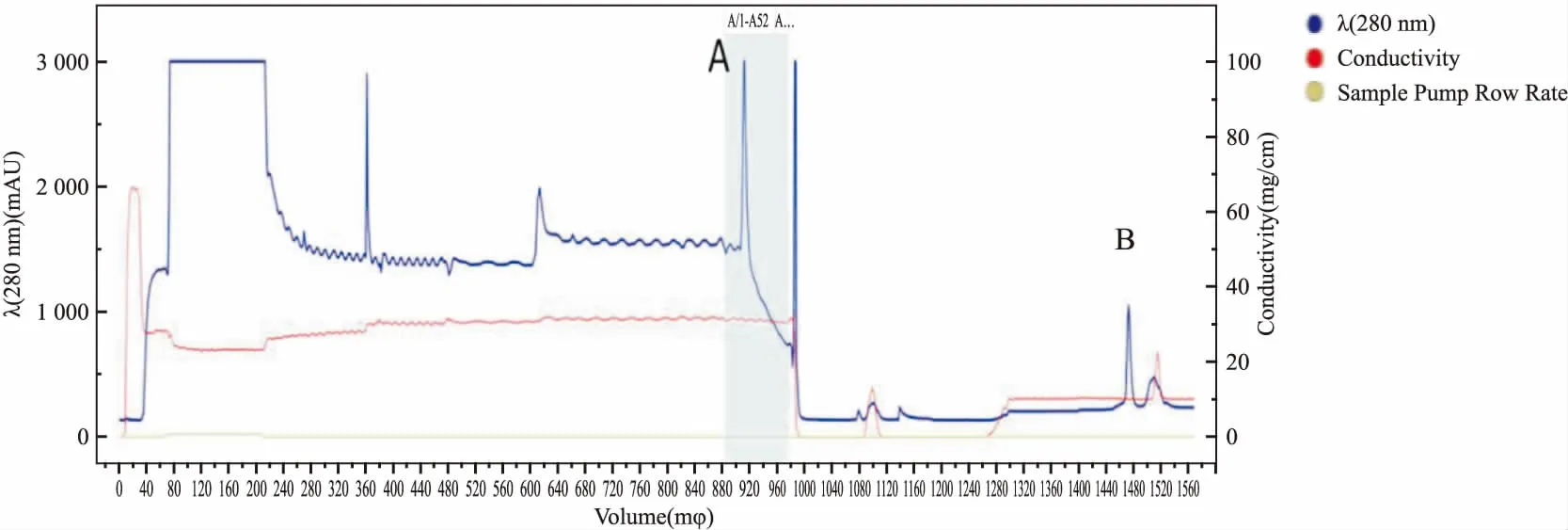
Fig 2 Purification of BLM642-1290 by Ni affinity chromatography and molecular sieve chromatography
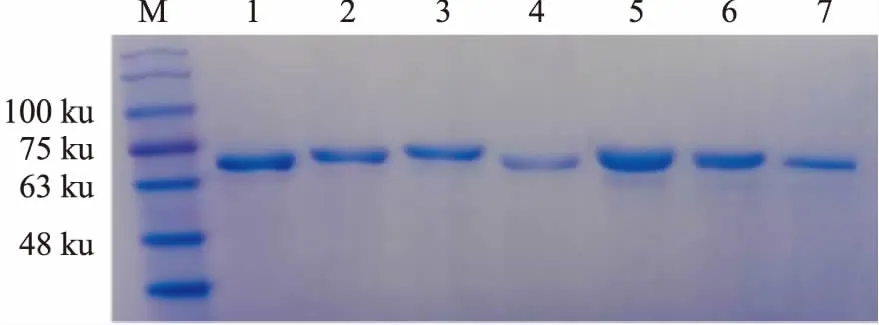
Fig 3 Determination of BLM helicase purity after molecular sieve chromatography
1.5 DMY对BLM642-1290解旋酶构象的影响缓冲液C中,终浓度为0.8 μmol·L-1BLM642-1290解旋酶与不同浓度梯度的DMY混合,25 ℃放置2 h后,于紫外分光光度计中扫描图谱,速度为每次3 min,该反应总体系为200 μL。另外,单独扫描不同浓度梯度的DMY在缓冲液C中的紫外吸收光谱。反应在室温下进行,扫描速度750 nm·min-1,扫描精度1 nm,扫描范围200~400 nm。每个图谱均为10~15个稳定数据的平均值。
1.6 DMY对BLM642-1290解旋酶二级结构的影响在1×PBS缓冲液中,终浓度为0.8 μmol·L-1BLM642-1290解旋酶与不同浓度的DMY混合,总反应体系为400 μL(1 mm光程比色皿),先室温下反应30 min后,再利用stop-flow电化学工作站(法国BioLogic Science Instruments公司)进行CD的检测,扫描速度每次5 min,直到CD图谱达到稳定,连续测定20次稳定后的CD图谱数据,拟合获得的20次实验数据,实验重复3次。色谱扫描区间为190~260 nm,扫描精度为1 nm,狭缝宽度为5 nm。
1.7 DMY对BLM642-1290解旋酶DNA结合活性的影响在此实验中,采用2种方式来研究DMY对BLM642-1290解旋酶DNA结合活性的影响[13]。第1种,BLM解旋酶与2 nmol·L-1的DNA(A2或A1A2)相互结合形成复合物,然后用不同浓度的DMY进行滴定;第2种,在室温下,不同浓度的DMY首先与BLM642-1290解旋酶反应5 min,然后再加入到2 nmol·L-1DNA(ssDNA或dsDNA)的反应缓冲液C中,荧光偏振仪检测。实验重复3次,数据为10~15个稳定偏振值平均之后的结果。
1.8 DMY对BLM642-1290解旋酶DNA解链活性的影响此方法借鉴刘金河等[13]的实验方法,25 ℃条件下,DMY与BLM642-1290解旋酶混合5 min后,加入到含2 nmol·L-1A1A2反应buffer中检测偏振值的变化,当偏振值达到稳定后,再加入终浓度为0.2 mmol·L-1ATP,10 s检测1次数据直到数据稳定,取数据稳定后的10~15个荧光偏振值数据求平均值,并进行3次实验重复。DNA解链的速率常数可根据方程(1)获得:
At=A1exp(-kobst)
(1)
式中At是解链时间为t时的荧光偏振值,A1是解旋酶与dsDNA完全结合时的荧光偏振值。
1.9 DMY对BLM642-1290解旋酶ATP酶活性的影响不同浓度DMY分别与终浓度10 nmol·L-1的BLM642-1290解旋酶混合5 min后,将该混合液与ATPase活性检测液按照1∶1(V/V)的比例混合,并开始计时,在2.5、5、7.5、10、15、20、25、30 min时间点,分别取出反应混合液200 μL,迅速加入到含50 μL Goldmix的EP管中,以终止ATP的水解反应,2 min后,加入稳定剂20 μL,平衡30 min,在650 nm波长下检测其吸光度值[13]。如果酶活力定义为每分钟催化1 mol底物水解所需的酶量(U·mL-1·min-1),BLM642-1290解旋酶的酶活力可由方程式(2)计算得出。
(2)
式中,A是根据标准曲线测出的Pi浓度(μmol·L-1);B是反应时间min;C是解旋酶的稀释倍数。
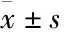
2 结果
2.1 DMY能够改变BLM642-1290解旋酶的空间结构DMY对BLM642-1290解旋酶的紫外吸收的影响如Fig 4所示,DMY在237 nm和330 nm处具有两个紫外吸收峰(Fig 4A),而BLM642-1290解旋酶在237 nm和276 nm处具有两个紫外吸收峰(Fig 4C)。当DMY与BLM642-1290解旋酶相互作用后,紫外吸收峰位发生了偏移,从237 nm处的红移到239 nm,峰型基本保持不变(Fig 4B)。进一步分析可知,在237 nm处,BLM642-1290解旋酶与DMY相互作用后的紫外吸收值大于BLM642-1290解旋酶与DMY的紫外吸收值之和(1.25>0.49+0.25)(Fig 4C),表明DMY与BLM642-1290解旋酶之间发生了相互作用,BLM642-1290解旋酶的构象发生了变化。将[DMY]/[BLM642-1290]的比值与紫外吸收值进行线性拟合,得一条直线(斜率k=0.0235),可知BLM642-1290解旋酶与DMY有且仅有1个结合位点(Fig 4D)。
2.2 DMY能部分解开BLM642-1290解旋酶二级结构由Fig 5A可知,BLM642-1290解旋酶的α-螺旋结构在不同浓度的DMY的作用下都有一定的结构变化,208 nm和222 nm处的负峰也清晰可见,表明BLM642-1290解旋酶的结构遭到一定程度的破坏,但是不能完全破坏其α-螺旋结构,而是存在一个最适DMY浓度。由Fig 5B可知,在DMY浓度为0~25 μmol·L-1时,随着DMY浓度的增加,α-螺旋在一定程度上逐渐解旋;当DMY浓度大于25 μmol·L-1时,随着DMY浓度的增加,α-螺旋又逐渐从松散状态基本回到最初的状态。
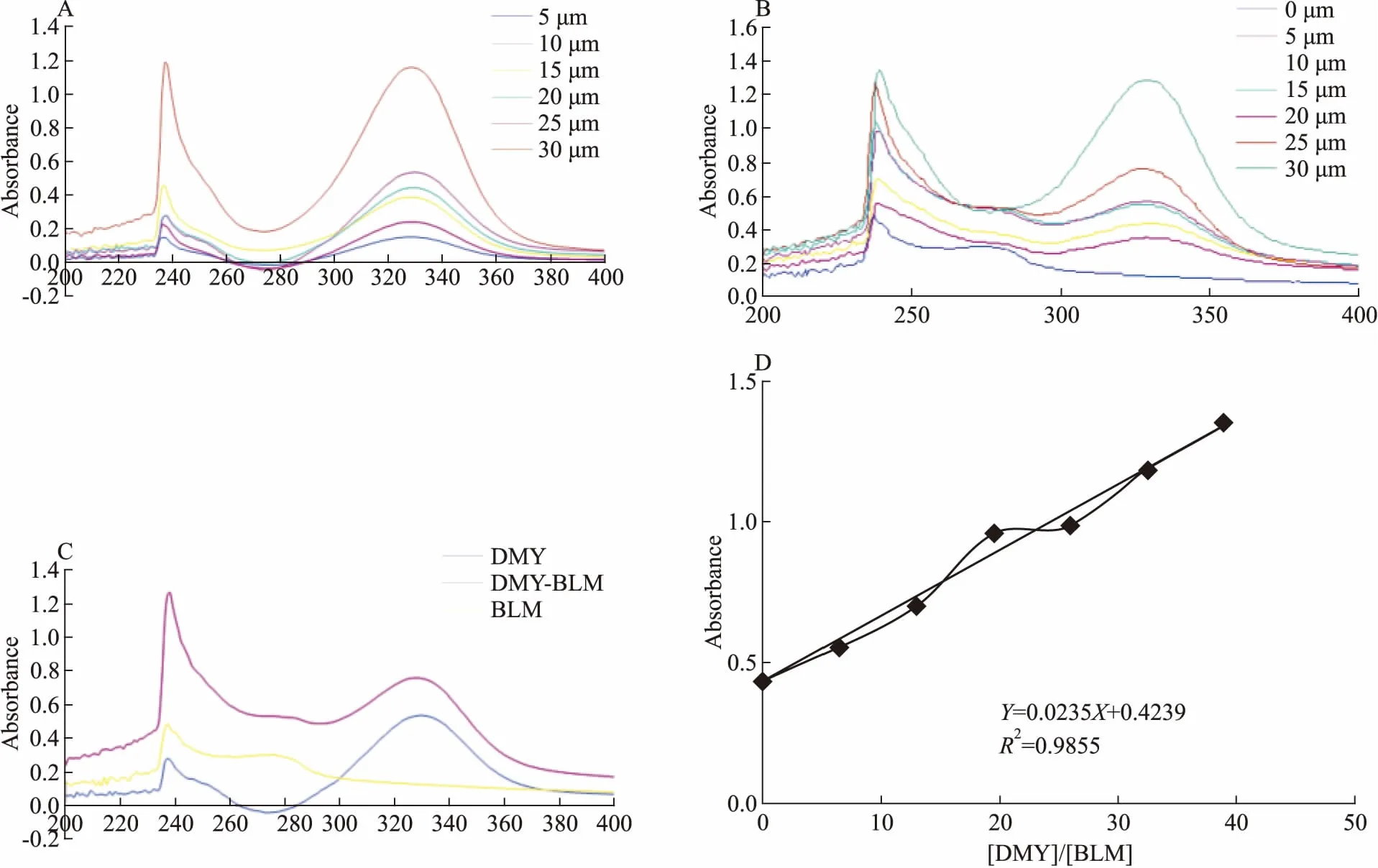
Fig 4 Ultraviolet(UV) absorption spectra of DMY interacted with BLM642-1290 helicase
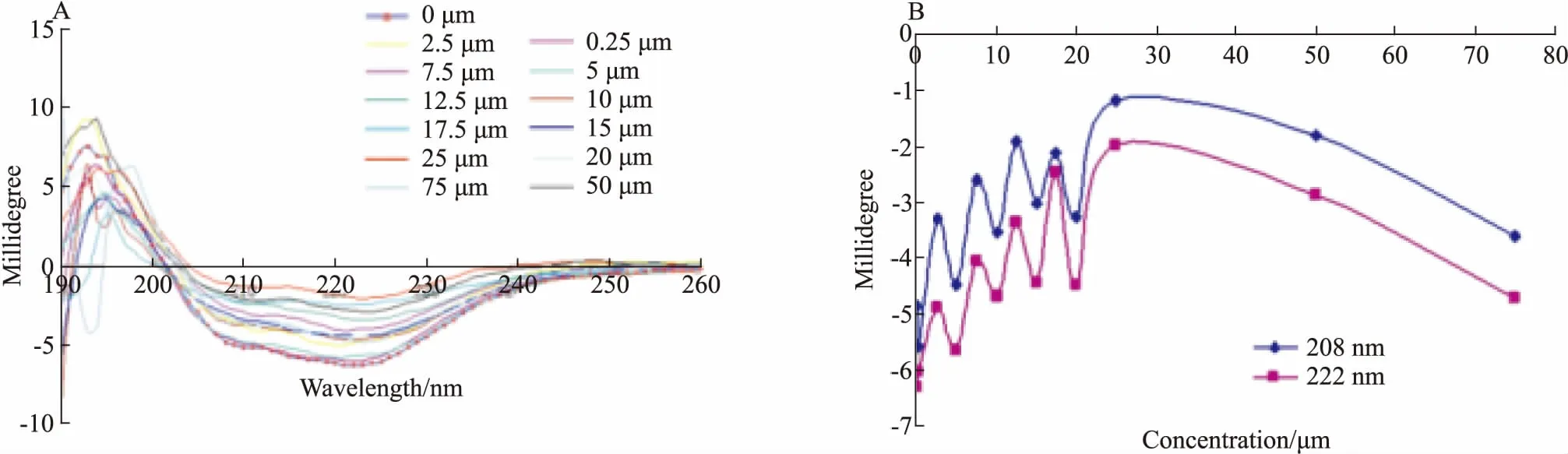
Fig 5 Circular dichroism(CD) spectra of DMY interacted with BLM642-1290 helicase
2.3 DMY能够抑制BLM642-1290解旋酶DNA结合活性DMY对BLM642-1290解旋酶的DNA结合活性的影响见Fig 6。由图可知,无论dsDNA(Fig 6A)还是ssDNA(Fig 6B)作为BLM642-1290解旋酶结合活性的底物,BLM642-1290与DNA相互结合而产生的荧光偏振值的增加量,都随DMY浓度的增加而明显减小(P<0.01);另外,DMY单独滴定dsDNA或者ssDNA的荧光偏振值无明显变化,说明DMY与DNA并无相互作用(Fig 6C)。由此可以推断,DMY与BLM642-1290解旋酶仅有的1个结合位点是在BLM642-1290解旋酶的DNA结合位点上,并且其与BLM642-1290解旋酶DNA结合位点的亲和力大于DNA。
2.4 DMY能抑制BLM642-1290解旋酶DNA解链活性DMY对BLM642-1290解旋酶的解链活性的影响见Fig 7。由Fig 7A可见,随着反应体系中DMY终浓度的逐渐增加(0~25.33 μmol·L-1),因解链而降低的荧光偏振值(A1-A2)逐渐减小,表明BLM642-1290解旋酶的解链活性逐渐降低。由此可知,BLM642-1290解旋酶的解链活性被DMY抑制,但不能完全抑制其解链。根据公式(1)进一步分析了DMY存在下BLM642-1290解旋酶的Kobs值的变化(Fig 7B),随着DMY浓度的增加,BLM642-1290解旋酶的Kobs值逐渐减小,表明DMY能够抑制BLM642-1290解旋酶的解链活性。
2.5 DMY能明显抑制BLM642-1290解旋酶的ATPase活性DMY对BLM642-1290解旋酶的ATPase活性的影响见Fig 8。由Fig 8A可知,DMY能明显抑制BLM642-1290解旋酶的ATPase活性(P<0.05),且酶活力的高低与DMY的浓度呈负相关,但不能完全抑制ATPase活性。由Fig 8B可知,在相同的酶浓度条件下,随着时间的延长,BLM642-1290解旋酶的酶活力逐渐下降,表明BLM642-1290解旋酶的酶活力与时间呈负相关。通过米氏方程双倒数作图法进一步分析实验数据,计算出酶反应的Vmax、Km等常数,结果见Tab 2。随着DMY浓度的增加,Vmax逐渐减小,而Km保持稳定,且Kcat减小,证明DMY竞争性抑制BLM解旋酶的ATPase活性。
3 讨论
BLM解旋酶是SF2超家族当中的一员,由解旋酶结构域(helicase)、RecQ保守的C末端结构域(RecQ-Ct)和解螺旋酶RNA酶D碳末端(HRDC)结构域组成[14]。Helicase结构域具有该家族特有的保守基序(Ⅰ、Ⅱa、Ⅲ-Ⅵ),结构和功能研究显示,motif Ⅰ和motif Ⅱ与NTP的结合有关。Motif Ⅳ和motif Ⅵ与ATP水解和核酸的结合有关,motif Q是SF2特有的保守序列,位于motif Ⅰ N末端前,其作用与ATP结合和水解有关。RecQ-Ct结构域具有DNA结合活性及ATPase活性,但不能结合ATP。HRDC结构域位于BLM解旋酶的C末端,能够辅助性结合DNA,进而调节其解链功能,也可增强和稳定BLM解旋酶与DNA的结合,但不具有催化活性[13]。
Fig 6 Effects of DMY on DNA-binding activity of BLM642-1290 helicase

Tab 2 Effects of DMY on ATPase activity constants of BLM642-1290 helicase
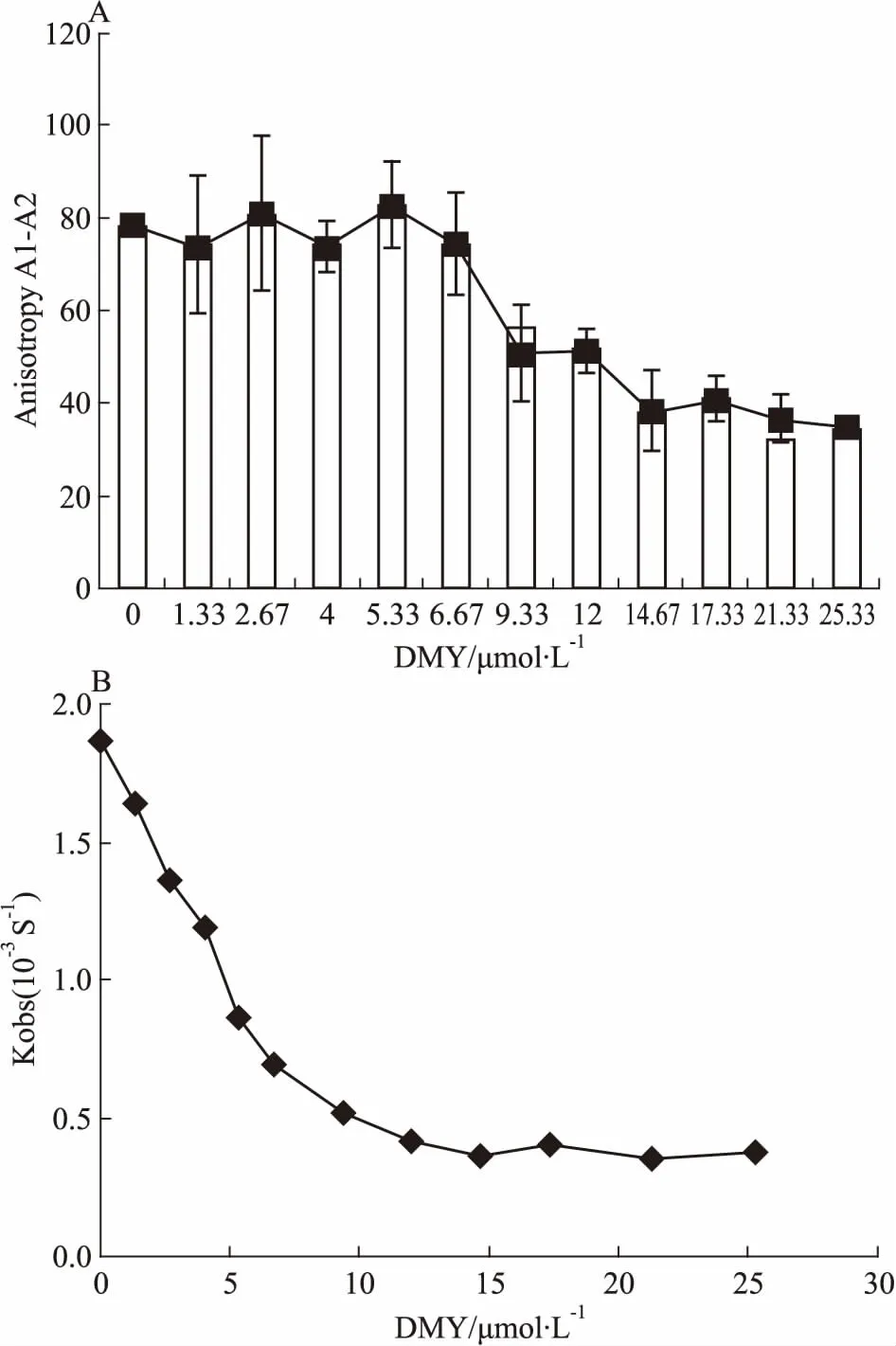
Fig 7 Effects of DMY on DNA unwinding activity of BLM642-1290 helicase
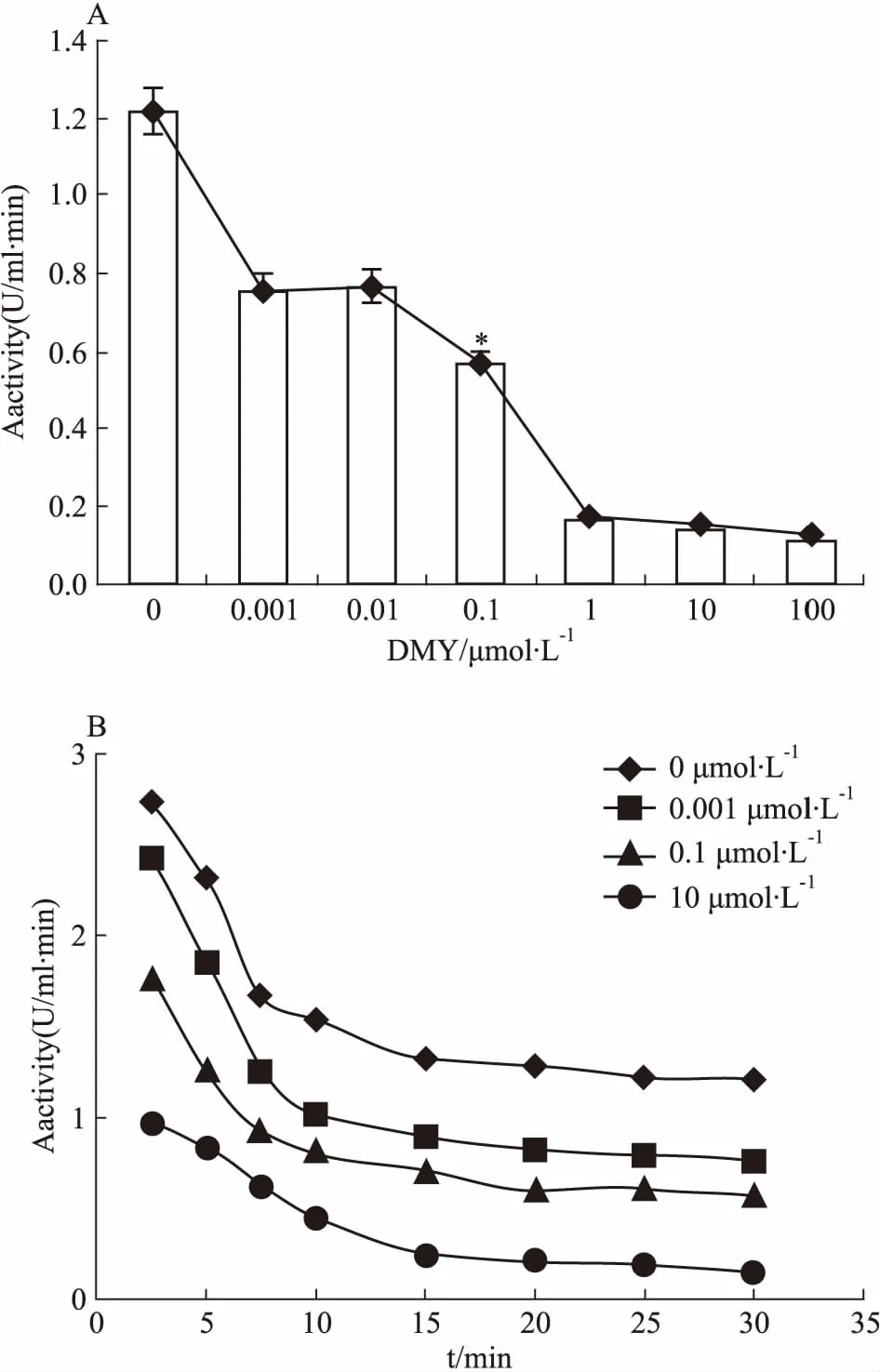
Fig 8 Effects of DMY on ATPase activity of BLM642-1290 helicase
从CD实验结果可知,DMY对BLM解旋酶的二级结构具有一定影响,使得BLM解旋酶的结构逐渐发生一定程度的松散,因α-螺旋结构的微变化,使得Helicase结构域内负责与DNA相互结合的活性基团的位置和距离发生微小变化,从而使得BLM与DNA的结合能力下降,而结合活性实验中进一步看到BLM解旋酶与DMY的结合力大于与DNA的结合能力,因此,DMY抢夺BLM解旋酶的DNA结合位点。从紫外光谱实验和解链活性实验可以看出,DMY与BLM解旋酶的结合仅有1个结合位点,而在结果“2.3”的DMY对BLM642-1290解旋酶DNA结合活性的影响当中可以看到,随着DMY的浓度逐渐增加,BLM-dsDNA或BLM-ssDNA的偏振值增加量(A1-A0)逐渐降低,甚至为零,也就是说DMY能将DNA从BLM642-1290置换下来,并占据DNA的结合位点,另外,因ATPase活性的实现区域在BLM642-1290解旋酶的helicase结构域中,为此我们大胆推断,这个结合位点很可能是在解旋酶结构域上,且阻止了motif Ⅳ和motif Ⅵ与DNA的结合,这种结合与DNA形成了可逆的竞争性机制。BLM解旋酶是人体细胞内普遍存在的一类具有解开双链DNA能力的活性蛋白质,它参与细胞内DNA的复制、重组、修复、转录等代谢过程,在维持染色体的稳定性方面具有重要的作用。BLM解旋酶在多种肿瘤细胞体内表达升高,比如乳腺癌、前列腺癌、肺癌、恶性淋巴瘤等[2,15],抑制BLM解旋酶的活性或表达量,能明显抑制肿瘤细胞的生长。例如,甘草次酸下调BLM解旋酶的表达,并抑制前列腺癌PC-3细胞的增殖[15],敲减BLM基因后的PC3细胞增殖能力、侵袭能力和迁移能力明显被丝裂霉素C抑制[16]。因此,寻找副作用小、价格便宜的小分子药物也就显得非常重要,这也为抗肿瘤药物的研发提供了新的思路。而提取DMY的植物在国内分布广泛、取材容易、成本低廉,成药后价格不高,值得进一步的研究。本实验的基础研究验证了DMY确实可以抑制癌症相关蛋白Bloom解旋酶的活性,也就在一定程度上说明DMY存在成为癌症治疗药物的可能性,后续的动物实验、临床实验的结果如何,有待进一步的研究。

Fig 9 Domain of BLM helicase
(致谢:本实验在贵州大学高原山地动物遗传育种与繁殖省部共建教育部重点实验室完成,衷心感谢各位同学和老师的支持和帮助)