Wnt10b与骨形态发生蛋白9诱导间充质干细胞成骨分化的关系
2019-09-24廖云鹏李福书孙文娟何百成
胡 莹,廖云鹏,李福书,周 娅,李 沁,孙文娟,何百成
(重庆市生物化学与分子药理学重点实验室,重庆医科大学药理学教研室,重庆 400016)
骨形态发生蛋白9(bone morphorgenetic protein 9,BMP9)作为BMP家族中研究较少的成员,是有效诱导间充质干细胞(mesenchymal stem cells,MSCs)骨向分化的因子之一[1]。因此,BMP9可能具有良好的临床应用前景,用于治疗骨质疏松、骨缺损、骨不连接等骨相关疾病。迄今为止,BMP9诱导MSCs骨向分化的具体分子机制仍不十分清楚。
作为TGF-β家族的一员,BMP9本身参与多种生理过程,如调节胆碱能神经元发育、铁代谢与血管生成,以及促进MSCs向成骨细胞系定向分化[2]等。BMP9可以通过经典BMP/Smad途径发挥其生理功能,此外,BMP9也可以通过非经典BMP/Smad途径发挥作用,如p38 MAPK和PI3K/Akt途径等[3]。研究报道,多种其他信号或因子能够参与调节BMP9诱导的MSCs成骨分化,如生长激素(growth hormone,GH)、胰岛素样生长因子(insulin-like growth factor,IGF)、维甲酸、Wnt/β-catenin及Notch信号[4]。BMP9虽能诱导MSCs骨向分化,但同时也伴有成脂分化[5]。因此,降低BMP9诱导MSCs的成脂分化可能是增强其诱导MSCs成骨分化的有效措施之一。
Wnt/β-catenin信号是调节细胞增殖分化的重要信号之一[6]。据报道,BMP9在诱导MSCs成骨分化的过程中能增强Wnt/β-catenin信号的活性,沉默β-catenin则减弱BMP9诱导MSCs成骨分化的能力[7]。但BMP9在诱导MSCs成骨分化过程中调节Wnt/β-catenin信号的机制尚不清楚。Wnt10b是Wnt/β-catenin信号的一种配体,对前脂肪细胞的分化有抑制作用,但能促进成骨分化[8]。因此,Wnt10b可能对BMP9诱导的MSCs成骨分化具有促进作用,尽管两者的关系目前还不清楚。本研究在前期工作基础上,分析Wnt10b在BMP9诱导MSCs成骨分化中的作用及其可能的分子机制。
1 材料与方法
1.1 材料
1.1.1实验动物 实验所用NIH孕鼠(E12.5),购自于重庆医科大学实验动物中心,并代养于SPF级饲养室内。
1.1.2细胞及细胞培养 小鼠胚胎成纤维细胞(mouse embryonic fibroblasts,MEFs),从NIH小鼠12.5 d胚胎中提取获得;人肾上皮细胞HEK293、间充质干细胞C3H10T1/2、成肌细胞C2C12及小鼠胚胎成骨细胞前体细胞MC3T3-E1,购自ATCC(Manassas,VA,USA)。采用高糖DMEM(含10%胎牛血清、100 kU·L-1青霉素、0.1 g·L-1链霉素)进行细胞培养,培养条件为5% CO2及37 ℃。
1.1.3试剂 一抗Wnt10b(ab155332)、环氧合酶2(cyclooxygenase-2,COX-2)(ab15191)、磷酸化环磷腺苷效应元件结合蛋白(phosphorylated cAMP-response element binding protein,p-CREB)(ab32096),购自Abcam公司;一抗GAPDH(sc-32233)、骨桥素(osteopontin,OPN)(sc-10593)、骨钙素(osteocalcin,OCN)(sc-18319)、重组人CCAAT增强子结合蛋白α(CCAAT/enhancer binding protein α,C/EBPα)(sc-61)、过氧化物酶体增殖物激活受体γ(peroxisome proliferator activated receptor γ,PPARγ)(sc-1984)、Smad1/5/8(sc-6031-R)、p-Smad1/5/8(sc-12353),购自Santa Cruz Biotechnology公司。
1.1.4仪器 细胞培养箱、低温离心机(Thermo公司);超净操作台(安泰技术有限公司);倒置荧光显微镜(日本Nikon);湿式转膜仪、垂直电泳槽(六一仪器厂);凝胶成像仪(Bio-Rad公司)。
1.2 重组腺病毒的构建及转染本实验所用重组腺病毒采用Ad-Easy系统进行构建[9]。利用PCR扩增BMP9、Wnt10b、绿色荧光蛋白(GFP)的编码序列,将所获片段以及Wnt10b的小干扰RNA片段克隆到腺病毒穿梭载体中,然后将穿梭质粒与AdEasy-1在BJ5183细菌中重组。最后,将重组好的质粒线性化,并转化到HEK293细胞中包装重组腺病毒。所获重组腺病毒载体分别命名为AdBMP9、AdWnt10b、AdGFP、AdsiWnt10b[10],其中,GFP用于标记腺病毒载体,AdGFP作为载体对照。
1.3 流式细胞术周期分析将C3H10T1/2细胞接种于6孔板中,待细胞贴壁后,根据实验设计要求,分别用不同的重组腺病毒(AdGFP、AdBMP9、AdWnt10b及AdBMP9合并AdWnt10b)处理细胞。48 h后收集细胞,用冰PBS(4 ℃)重悬细胞并离心(800 r·min-1, 5 min×2次)。然后用预冷的70%、50%、30%乙醇固定,并用PBS洗涤。加入1 mL PI(20 g·L-1)溶液(含RNase,1 g·L-1)孵育30 min。最后用流式细胞仪进行周期分析。每组实验重复3次。
1.4 Real-time PCR实验利用TRIzol试剂裂解细胞并提取总RNA,通过逆转录反应得到cDNA。将所得cDNA稀释5~10倍,作为模板用于定量PCR检测。以GAPDH的表达水平对目的基因的表达进行标准化。本实验所涉及到引物序列见Tab 1。
1.5 Western blot实验将细胞接种于6孔板中,根据实验设计利用重组腺病毒以及其他试剂处理。在预定的时间点,裂解细胞,并将裂解液煮沸10 min使裂解物变性。所得样品在SDS-PAGE胶中分离,并转移到PVDF膜上,以10%脱脂奶粉封闭,以相应的一抗和二抗分别孵育。最后利用超敏ECL化学发光试剂测定靶蛋白表达情况。每组实验重复3次。
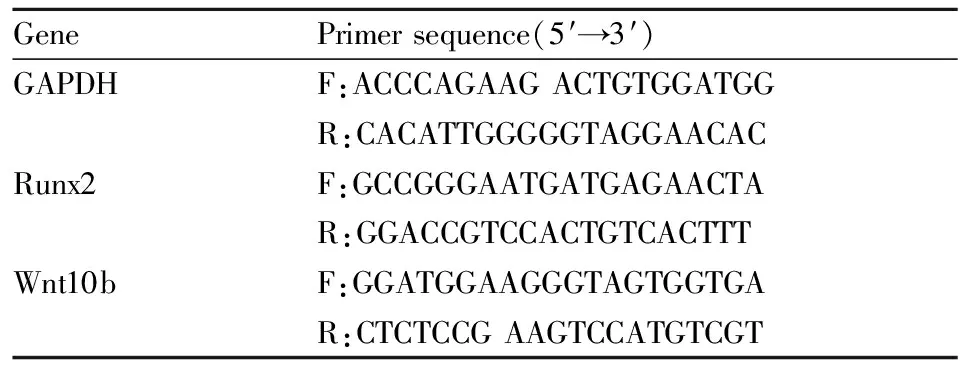
Tab 1 Primer sequences used for real-time PCR
1.6 矿物质沉积实验将细胞接种于24孔板中,并根据实验设计用相应重组腺病毒及其他试剂处理。21 d后,弃培养基并用PBS洗涤3次,室温下加入0.05%戊二醛固定10 min,用蒸馏水洗涤。然后以0.4%茜素红S溶液对细胞进行染色5 min,用蒸馏水轻轻洗涤。观察矿物质沉淀。扫描平板,并在显微镜下采集图像。每组实验重复3次。
1.7 油红O染色将细胞接种到24孔板中,根据实验设计用重组腺病毒及其他相应试剂处理细胞。14 d后,先用10%福尔马林固定细胞30 min,然后弃固定液并用PBS洗涤3次,最后用油红O溶液染色,在显微镜下采集图像。定量分析时,用异丙醇从细胞中提取油红O,在570 nm处测定吸光度值。每组实验重复3次。
2 结果
2.1 BMP9在干细胞中对Wnt10b表达的影响Wnt10b是经典Wnt/β-cantein信号的一种配体。课题组前期研究表明,BMP9能够促进MSCs的成骨分化,但BMP9与Wnt10的关系尚不清楚。因此,本研究旨在探讨Wnt10b是否参与调节BMP9诱导MSCs成骨分化的作用。首先检测在MSCs中,BMP9对Wnt10b表达的影响。结果显示,BMP9在C3H10T1/2细胞及3T3-L1细胞中能上调Wnt10b的mRNA水平(Fig 1A、1B),并且增加C3H10T1/2细胞中Wnt10b的蛋白表达水平(Fig 1C)。此外,Wnt10b在C3H10T1/2、C2C12、MEFs和MC3T3-E1等细胞中均有表达(Fig 1D)。以上结果提示,Wnt10b可能在BMP9诱导MSCs成骨分化过程中起重要调节作用。
2.2 Wnt10b对BMP9诱导C3H10T1/2细胞成骨分化的影响BMP9可在C3H10T1/2细胞中上调Wnt10b表达。因此,本研究接下来分析Wnt10b是否可以增强BMP9诱导C3H10T1/2细胞成骨分化的能力。定量PCR检测结果显示,BMP9能够上调Dlx-5 mRNA的表达水平,Wnt10b也能增加Dlx-5 mRNA的表达水平,并明显增强BMP9上调Dlx-5表达的能力(Fig 2A)。Western blot结果显示,BMP9增加C3H10T1/2细胞中OPN和OCN的蛋白水平,Wnt10b对OPN和OCN的蛋白水平没有明显影响,但能明显增强BMP9对OPN和OCN表达的促进作用(Fig 2B)。同时,Wnt10b还能明显增强BMP9诱导MSCs矿物质沉积的能力(Fig 2C)。沉默Wnt10b不仅降低Dlx-5 mRNA水平,也减弱BMP9上调Dlx-5表达的能力(Fig 2D),沉默Wnt10b也减弱BMP9上调OPN和OCN蛋白水平的作用(Fig 2E)。矿化实验结果也显示,沉默Wnt10b能减弱BMP9在C3H10T1/2细胞中矿化的能力(Fig 2F)。以上结果提示,Wnt10b可能是促进BMP9诱导MSCs成骨分化的重要调节因子。
2.3 BMP9及Wnt10b对C3H10T1/2细胞增殖的影响为分析Wnt10b促进BMP9诱导干细胞骨向分化的机制,本研究分析了Wnt10b与BMP9对细胞周期的影响。流式分析结果显示,BMP9及Wnt10b均不影响细胞的周期(Fig 3A)。 Western blot分析也显示,BMP9及Wnt10b均不明显影响PCNA的蛋白水平(Fig 3B)。以上结果提示,Wnt10b促进BMP9诱导C3H10T1/2细胞成骨分化可能与影响增殖能力无关。

Fig 1 Effect of BMP9 on expression of Wnt10b in stem cells

Fig 2 Effect of Wnt10b on BMP9-induced osteogenic markers in C3H10T1/2 cells

Fig 3 Effect of BMP9 and Wnt10b on cell cycle in C3H10T1/2 cells
2.4 Wnt10b对BMP9介导的BMP/Smad信号转导的影响BMP9可通过经典BMP/Smad途径或非经典BMP/Smad途径发挥其生理功能。因此,本研究进一步检测Wnt10b促进BMP9诱导干细胞成骨分化,是否与影响BMP/Smad信号的活性有关。Western blot结果显示,BMP9对Smad1/5/8水平无明显影响,但能明显增加Smad1/5/8的磷酸化水平。Wnt10b对p-Smad1/5/8和Smad1/5/8的水平无明显影响,但能明显增强BMP9对Smad1/5/8磷酸化的促进作用(Fig 4A、4B)。沉默Wnt10b对p-Smad1/5/8和Smad1/5/8的水平也无明显影响,但能明显降低BMP9对Smad1/5/8磷酸化的促进作用(Fig 4C、4D)。以上结果提示,Wnt10b促进BMP9诱导干细胞成骨分化的作用可能与增强BMP/Smad信号的转导活性有关。
2.5 Wnt10b对BMP9诱导C3H10T1/2细胞成脂分化的影响干细胞的成骨分化和成脂分化是两个相互影响的生理过程,脂肪生成增加可能导致骨形成减少。据报道,Wnt10b可以抑制前脂肪细胞分化。因此,我们推测Wnt10b也可能通过抑制BMP9介导的MSCs成脂分化,从而增强BMP9的成骨分化诱导能力。油红O染色结果显示,Wnt10b明显降低BMP9诱导C3H10T1/2细胞成脂分化的能力(Fig 5A、5B)。Western blot结果显示,BMP9上调C/EBPα和PPARγ的蛋白水平,但Wnt10b能明显减弱BMP9增加C/EBPα和PPARγ水平的作用(Fig 5C)。沉默Wnt10b能促进干细胞向脂肪细胞分化,同时明显增强BMP9诱导的脂滴形成(Fig 5D、5E),并且增强BMP9上调C/EBPα和PPARγ蛋白水平的能力(Fig 5F)。以上结果提示,Wnt10b对BMP9诱导干细胞成骨分化的作用可能部分与抑制BMP9诱导干细胞成脂分化有关,但具体机制有待进一步深入研究。

Fig 4 Effect of Wnt10b on BMP9-induced BMP/Smad signaling n=3)

Fig 5 Effect of Wnt10b on BMP9-induced adipogenic differentiation of C3H10T1/2 cells
3 讨论
本研究分析了Wnt10b对BMP9诱导C3H10T1/2细胞成骨分化的影响,以及调节该过程的可能分子机制,首次提出Wnt10b增强BMP9诱导成骨分化的能力可能与增强BMP/Smad信号的转导活性有关,并且进一步探索了其对BMP9成脂诱导分化能力的影响,为进一步揭示Wnt信号通路与BMP/Smad通路之间的关系提供实验基础。结果发现,BMP9能上调Wnt10b的表达,Wnt10b能明显促进BMP9诱导C3H10T1/2细胞成骨分化的能力;Wnt10b的这种作用可能与增强BMP/Smad信号的传导,同时抑制BMP9诱导干细胞的成脂分化有关。
BMP属于转化生长因子β(TGF-β)超家族成员,是调节细胞多种重要生理过程的关键因子,如细胞的增殖和分化[4]。虽然BMP9(也称为GDF2)最开始是从小鼠肝脏的cDNA文库中鉴定,但其具有多种生理功能,包括调节胆碱能神经元的发育、铁代谢、血管生成,以及促使MSCs向成骨细胞系的定向分化等[1-2]。据报道,BMP9的成骨分化诱导能力可能强于BMP2和BMP7。因此,BMP9可能是最有效的诱导干细胞成骨分化的BMPs因子之一[1]。BMP9通常通过BMP/Smad途径(即经典BMP/Smad途径)或非经典BMP/Smad途径(如p38 MAPKs和PI3K/Akt途径)发挥其生理功能[3]。在经典的BMP/Smad途径中,BMP9与其膜受体结合,并激活Smad1/5/8(即p-Smad1/5/8)。然后,p-Smad1/5/8募集并磷酸化Smad4以形成复合物。最后,复合物易位到细胞核中,调节下游靶基因表达。此外,有研究表明,多种其他因子或信号也参与调节BMP9诱导的干细胞成骨分化,如GH、IGF、维甲酸和Wnt/β-catenin信号[3-4]。但BMP9诱导干细胞成骨分化的具体分子机制尚不十分清楚。
成骨和成脂分化是两个相互影响的过程[11]。MSCs通常以成脂分化的减少为代价,向成骨细胞谱系分化,反之亦然。尽管BMP9在MSCs中显示出强大的诱导成骨分化的能力,但它也可诱导干细胞向脂肪细胞分化[12]。因此,可以通过减弱MSCs中的成脂分化,来增强BMP9的促成骨能力。Wnt/β-catenin是调节细胞分化、增殖和存活的重要信号。研究表明,Wnt信号对骨骼系统和脂肪的形成也非常重要[13]。异常的Wnt/β-catenin信号与多种骨相关疾病有关,如骨质疏松症、骨关节炎、骨硬化症等。据报道,BMP9在诱导MSCs成骨分化时可激活Wnt/β-catenin信号,沉默β-catenin或过表达DKK1均能降低BMP9的促成骨潜能,而过表达β-catenin则增强BMP9的促成骨能力[14]。因此,BMP9诱导干细胞成骨分化作用与Wnt/β-catenin信号关系密切。
Wnt10b是Wnt/β-catenin信号的另一种经典蛋白质。有研究表明,Wnt10b与免疫系统的调节有关[15]。此外,Wnt10b可以促进成骨分化,并抑制前脂肪细胞的分化。因此,课题组推测,Wnt10b可能参与调节BMP9诱导的成骨分化。本研究结果显示,BMP9可以上调C3H10T1/2细胞和其他几种干细胞中Wnt10b的表达;Wnt10b能增加BMP9诱导的MSCs成骨指标水平,但沉默Wnt10b则降低BMP9的这种作用。提示Wnt10b对BMP9诱导MSCs的成骨分化具有重要促进作用。研究进一步发现,Wnt10b明显增加由BMP9介导的Smad1/5/8磷酸化水平,而沉默Wnt10b减弱BMP9对p-Smad1/5/8的促进作用。同时,Wnt10b可降低BMP9在C3H10T1/2细胞中上调PPARγ和C/EBPα水平的作用,并减少细胞中脂滴的形成;沉默Wnt10b增强BMP9在C3H10T1/2细胞中诱导的脂肪分化。
综上所述,本研究的结果表明,Wnt10b可以促进BMP9诱导MSCs的成骨分化,这种作用可能与增强BMP/Smad信号的传导活性,以及抑制BMP9诱导MSCs成脂分化有关。
[致谢:本研究在重庆市生物化学与分子药理学重点实验室完成,感谢何通川教授(T. C. HE,芝加哥大学医学中心)为本实验馈赠所需的重组腺病毒载体。]