大麦β-淀粉酶基因在大肠杆菌中的异源表达
2016-07-21朱培李崎朱林江贺鹏宇李永仙
朱培,李崎,朱林江,贺鹏宇,李永仙*
1(江南大学 酿酒科学与工程研究室,江苏 无锡,214122) 2(江南大学 生物工程学院,江苏 无锡,214122)
大麦β-淀粉酶基因在大肠杆菌中的异源表达
朱培1,李崎2,朱林江3,贺鹏宇4,李永仙1*
1(江南大学 酿酒科学与工程研究室,江苏 无锡,214122)2(江南大学 生物工程学院,江苏 无锡,214122)
摘要大麦来源的β-淀粉酶具有稳定性高和工业应用效果好的优点,被广泛应用于食品、酿造以及粮食加工,但是其制备成本高,价格较昂贵。该研究使用大肠杆菌异源表达大麦来源的β-淀粉酶。首先通过基因合成技术获得密码子优化的大麦来源的β-淀粉酶基因,将该基因通过质粒pET28a(+)在大肠杆菌BL21(DE3)中过量表达获得重组β-淀粉酶。其次,对重组表达的β-淀粉酶和大麦中直接提取的β-淀粉酶进行酶学性质比较分析。结果表明,重组表达的β-淀粉酶其分子质量大小与大麦中β-淀粉酶一致,即59 kDa,重组的β-淀粉酶比酶活由大麦来源的588 U/mg降为285.5 U/mg,最适作用温度降低了10 ℃,最适pH保持一致。所以,大麦β-淀粉酶能够在细菌中高效表达,但是其酶学性质发生较大改变。
关键词大麦;β-淀粉酶;异源表达;大肠杆菌
淀粉酶是水解淀粉和糖原的一类酶的总称,主要存在于动物、植物以及微生物中[1],一般作用于可溶性淀粉、糖原、α-1,4-葡聚糖等,水解其中的α-1,4-糖苷键。根据氨基酸序列的同源性,分属糖苷水解酶类的14族[2]。根据淀粉酶对淀粉的不同催化水解方式,可将其分为4种淀粉酶,即α-淀粉酶、β-淀粉酶、葡萄糖淀粉酶和脱支酶。其中β-淀粉酶,又称α-1,4-葡聚糖麦芽糖水解酶(EC3.2.1.2),是一种外切型淀粉酶。它从淀粉的非还原性末端开始,依次切开间隔的α-1,4糖苷键,生成麦芽糖和β-极限糊精[3],其在啤酒酿造、糖浆制造、纺织与医疗中具有重要应用[4-5]。
现有β-淀粉酶酶制剂主要来源于植物提取和微生物发酵生产。其中植物来源β-淀粉酶酶制主要提取于大麦、甘薯、大豆等粮食作物中[6],而微生物生产β-淀粉酶酶制主要是通过枯草芽孢杆菌等表达生产。相比于微生物来源的β-淀粉酶,植物来源的β-淀粉酶具有酶活高、热稳定性好、pH作用范围广等特点[7],所以受工业应用的偏爱,其中应用最广泛的是大麦来源的β-淀粉酶酶制剂。但是,大麦β-淀粉酶经过提取获得,不但消耗大量的大麦[8],而且制备成本偏高,其价格高居不下。虽然微生物产β-淀粉酶可实现高酶活发酵,而且诱变后可提高其酶活力和热稳定性[9],但是根据工厂应用经验,比如在制糖过程中,微生物产β-淀粉酶仍难以达到大麦来源酶制剂的使用效果。因此本论文将大麦β-淀粉酶基因导入大肠杆菌中进行异源表达[8,10],研究大肠杆菌重组的β-淀粉酶的性质,同时将其与大麦来源的β-淀粉酶进行比较。
1材料与方法
1.1材料
1.1.1β-淀粉酶基因来源
通过NCBI查得大麦来源β-淀粉酶基因序列(GenBank:D21349.1),交付苏州金唯智生物科技有限公司合成,获得完整的密码子优化的大麦β-淀粉酶基因序列,预设计酶切位点为XhoI与NdeI。
1.1.2质粒以及其他试剂
质粒pMD19T-simple购自TaKaRa生物工程有限公司,表达载体pET28a(+)质粒,大肠杆菌JM109与BL21(DE3)均由本研究室保藏。TaqDNA polymerase、克隆载体pMD19T-simple、Mini BEST基因组提取试剂盒、感受态细胞制作试剂盒、T4DNA连接酶、限制性内切酶XhoI和NdeI购自TaKaRa生物工程有限公司;San prep柱式DNA胶回收试剂盒购自上海生工生物工程有限公司。
1.2方法
1.2.1重组表达载体的构建
大麦β-淀粉酶基因amyB首先插入到pMD19T-simple载体进行亚克隆以方便基因后续插入到pET28a(+)中,将重组质粒交由上海生工公司测序。双酶切获得目的基因,将目的基因与双酶切后的pET28a(+)质粒连接后,涂布与含硫酸卡那霉素的LB平板于37 ℃培养过夜,挑选阳性pET28a(+)-amyB重组表达质粒单菌落于50 mL LB培养基后,提取质粒进行双酶切验证。所有限制性酶切反应体系均参照试剂盒说明书配制。
1.2.2大肠杆菌的诱导表达
挑取含pET28a(+)-amyB重组表达质粒单菌落于含有硫酸卡纳霉素的LB液体培养基中,37 ℃ 200 r/min过夜培养;以4%的接种量转接到含硫酸卡纳霉素的50 mL的新鲜LB液体培养基中,37 ℃ 200 r/min培养至OD600≈1.0时加入诱导剂IPTG至终浓度为0.06 mmol/L;转移至24 ℃ 150 r/min培养6 h后于4 ℃ 8 000 r/min离心10 min,收集菌体及上清液;上清液经透析后使用蛋白超滤管浓缩,于4 ℃保存,同时培养含有空pET28a(+)质粒的宿主菌,诱导培养作为阴性对照。
1.2.3重组蛋白的收集与纯化鉴定[11-12]
菌液离心去其上清液,加入磷酸缓冲液悬浮后在冰浴条件下利用超声细胞破碎仪破碎细胞,使超声时间累积15 min(工作时间:4 s;间隙时间:6 s;工作次数:90)后4 ℃ 8 000 r/min离心10 min,收集上清液,作为粗蛋白样品。用pH 7.0磷酸盐缓冲液稀释10倍后,采用镍柱纯化和AKTA蛋白纯化系统进行分离,其中AKTA蛋白纯化选择阴离子交换色谱Mono Q。纯化结束后收集吸收峰,进行SDS-PAGE电泳检测。
1.2.4大麦来源β-淀粉酶的纯化
用pH 6.0磷酸缓冲液稀释大麦来源的β-淀粉酶酶制剂,采用AKTA蛋白纯化系统进行分离,选择凝胶柱Superdex G75。过柱完成后收集吸收峰,经过SDS-PAGE电泳进行检测。
1.2.5β-淀粉酶的酶学性质分析[13-14]
使用Megazyme公司的Beta-Amylase Assay Procedure试剂盒分别测定β-淀粉酶的最适温度(温度范围40~70 ℃)、50 ℃温度稳定性、最适pH(pH范围4.0~8.0)与pH 6.0时的稳定性。
2结果与分析
2.1pET28a(+)-amyB表达载体的构建与在大肠杆菌中的诱导表达
pET28a(+)-amyB的XhoI与NdeI双酶切结果如图1A所示,图中大小在1 600 bp左右为目的基因,测序结果证实重组质粒构建正确。将其重组质粒转化到大肠杆菌BL21(DE 3)中进行诱导表达。诱导表达后,收集破碎细菌液上清后经SDS-PAGE分析,由图1B可见,与诱导表达空pET28a(+)质粒的宿主菌相比较,重组载体诱导表达后在60 kDa左右处有明显的特异条带,说明新载体在诱导表达后有新蛋白产生,而大麦来源的β-淀粉酶的蛋白分子质量在59 kDa左右。此外,对该蛋白条带进行回收纯化,用于质谱鉴定,其肽段匹配结果与蛋白NCBI ID为gi|113786相似度最大,分子质量为59 609,为大麦β-淀粉酶来源。
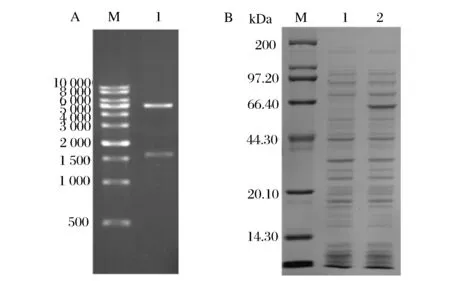
(A)重组质粒pET28a(+)-amyB的XhoI与NdeI双酶切核酸电泳图M-DNAMarker;1-XhoI和NdeI双酶切)(B)重组质粒pET28a(+)-amyB大肠杆菌中的诱导表达β-淀粉酶电泳图M-蛋白质标准分子量;1-诱导含空质粒pET28a(+)的大肠杆菌胞内蛋白;2-诱导含质粒pET28a(+)-amyB的胞内蛋白图1 重组质粒pET28a(+)-amyB的双酶切验证与重组β-淀粉酶的诱导表达电泳图Fig.1 Digestion of recombinant plasmid and induced expression of recombinant β-amylase
2.2重组β-淀粉酶的纯化[15]
由于重组β-淀粉酶含有组氨酸标记,因此首选镍柱纯化粗酶液。多次重复试验表明目的蛋白与杂蛋白同时被洗脱,重组β-淀粉酶无法与镍柱结合。如图2A所示,目的蛋白随着杂蛋白在50 mmol/L咪唑浓度下一起被洗脱下来(箭头所示为目的蛋白)。因此改用AKTA蛋白纯化系统来纯化重组表达的β-淀粉酶。收集的上清液经过透析,超滤浓缩后,选择阴离子交换色谱柱Mono Q进一步纯化β-淀粉酶,通过预实验,采用含有0.15 mol/L NaCl pH 7.4的磷酸盐缓冲液洗脱,收集洗脱峰经过SDS-PAGE鉴定其中峰A6是BL21(DE3)异源表达的大麦β-淀粉酶,如图2C所示,纯化的最终结果得到1条单一的蛋白条带,大小在59 kDa左右。
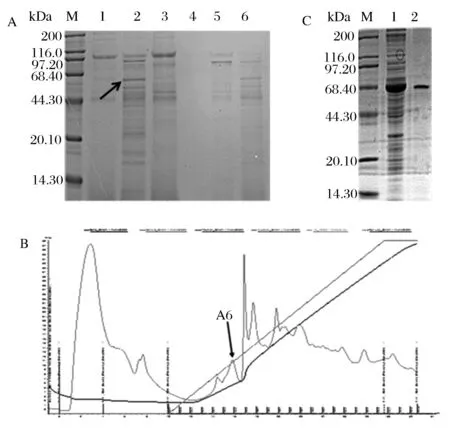
(A)粗酶液的镍柱洗脱结果的SDS-PAGE电泳比较M-蛋白质标准分子量;1~6-洗脱液咪唑浓度分别为:20、50、100、150、200、250 mmol/L(B)粗酶液的MonoQ纯化峰图(C)重组β-淀粉酶的诱导表达电泳图M-蛋白质标准分子量;1-纯化前的重组蛋白;2-纯化后的重组蛋白图2 重组菌诱导表达β组淀粉酶的纯化Fig.2 Purification of expressed β-amylase
2.3大麦来源的β-淀粉酶的纯化
为了比较分析大肠杆菌中异源表达的β-淀粉酶性质,对同一大麦品种来源的β-淀粉酶制剂进行了纯化。使用AKTA蛋白纯化系统纯化大麦来源β-淀粉酶酶制剂,选择凝胶色谱柱Superdex G75纯化大麦β-淀粉酶,以pH 6.0的PBS洗脱。凝胶过滤色谱Superdex G75峰图见图3A,SDS-PAGE鉴定结果为峰B6为β-淀粉酶,纯化结果如图3B所示,SDS-PAGE得到单一清晰条带,蛋白分子质量在59 kDa附近。
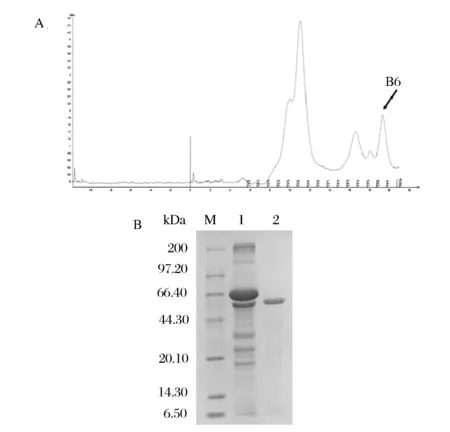
(A)β-淀粉酶Superdex G-75纯化峰图(B)大麦来源β-淀粉酶的SDS-PAGE结果M-蛋白质标准分子量;1-纯化前的粗蛋白;2-纯化后的β-淀粉酶图3 大麦来源β-淀粉酶的纯化Fig.3 Purification of β-amylase from Barley
2.4重组β-淀粉酶与大麦来源β-淀粉酶的酶学性质比较
取纯化后的重组β-淀粉酶酶液(485.5 U/mL)与大麦来源纯化的β-淀粉酶酶液(10 000 U/mL),使用爱尔兰Megazyme公司的Beta-Amylase Assay Procedure试剂盒专一性测定β-淀粉酶的酶活,从而比较比酶活、最适温度、温度稳定性、最适pH与pH稳定性,结果如表1所示。

表1 不同来源β-淀粉酶酶学性质的比较
如图4A所示,大麦来源的β-淀粉酶的最适温度为55 ℃,大肠杆菌重组表达的β-淀粉酶的最适温度变为45 ℃,与大麦来源β-淀粉酶相比,重组酶的最适温度下降了10 ℃。
分别在50 ℃条件下处理大麦来源、大肠杆菌表达的β-淀粉酶,测其酶活,结果如图4B所示。在50 ℃时,重组蛋白的半衰期相比原始β-淀粉酶减少了40 min,发现β-淀粉酶在大肠杆菌中重组表达后热稳定性显著下降。
使用不同pH的磷酸缓冲液比较β-淀粉酶酶活,在40 ℃条件下测其酶活,如图4C所示,两种β-淀粉酶的最适pH均为6.0。
在pH 6.0的磷酸缓冲液中,在40 ℃条件下测两种β-淀粉酶的pH稳定性,如图4D所示。两种β-淀粉酶的半衰期大概都在6 h左右,下降趋势差别不大,经过大肠杆菌的重组表达后,β-淀粉酶的pH稳定性与原大麦β-淀粉酶的pH稳定性相比基本保持不变。

(A)最适温度的比较;(B)50 ℃时的热稳定性的比较;(C)最适作用pH的比较;(D)pH 6.0时的稳定性的比较图4 重组β-淀粉酶与直接来源大麦的β-淀粉酶的酶学性质比较分析Fig.4 Comparative analysis of enzymatic properties between recombinant β-amylase and purified barley β-amylase
3讨论
论文针对大麦来源的β-淀粉酶在大肠杆菌中异源表达进行了研究,结果表明经过密码子优化的大麦β-淀粉酶能够在大肠杆菌中高效表达,通过SDS-PAGE分析能够清晰判断,高于之前的研究报道[16]。但是,经过凝胶色谱柱和离子交换柱纯化后的重组β-淀粉酶酶学性质与大麦中直接提取的β-淀粉酶具有一定差异。重组β-淀粉酶的热稳定性下降了10 ℃,比酶活由大麦来源的588 U/mg降为285.5 U/mg;而重组表达对酶的最适作用pH和pH稳定性影响不大。重组的β-淀粉酶的热稳定性和比酶活下降,这一研究结果与之前的报道不一致,可能是由于不同的β-淀粉酶基因、不同的E.coli菌株、不同的酶纯化方法造成的。而本次研究重组β-淀粉酶的热稳定性和比酶活均低于原大麦中纯化的酶,这可能是由于真核来源的蛋白质在原核生物宿主中表达时,其三级结构的折叠以及蛋白的修饰不同造成的。大麦β-淀粉酶是来源于真核生物,通过糖基化修饰后可使其结构更加稳定[17],而大肠杆菌的表达系统并无糖基化修饰系统,糖基化或许是重组蛋白最适温度与温度稳定性下降的原因。
此外,对纯β-淀粉酶的研究过程中发现,不管是重组表达的β-淀粉酶,还是直接从大麦中纯化的β-淀粉酶,都是一种极不稳定的酶,容易失活。然而在自然状态下,大麦提取的β-淀粉酶酶制剂可以较稳定的保持活性。因此,通过本次纯的β-淀粉酶的比较研究,推测在大麦提取液中存在某些物质,能够保护β-淀粉酶,提高其酶活半衰期,这也可能是植物来源的β-淀粉酶酶制剂具有更好地工业应用效果的原因之一。
参考文献
[1]邬显章. 酶的工业生产技术[J]. 长春: 吉林科学技术出版桂, 1988: 40-41.
[2]Henrissat B, Davies G. Structural and sequence-based classification of glycoside hydrolases[J]. Current opinion in structural biology, 1997, 7(5): 637-644.
[3]王海明. β-淀粉酶与高辅料啤酒酿造[J]. 啤酒科技, 2000(6): 6-9.
[4]齐继艳, 陈舟舟, 卢晗, 等. 植物 β-淀粉酶[J]. 植物生理学通讯, 2008, 44(2): 334-340.
[5]孙俊良, 梁新红, 贾彦杰, 等. 植物 β-淀粉酶研究进展[J]. 河南科技学院学报:自然科学版,2011,39(6):1-4.
[6]Ventanas J, Córdoba J, Antequera T, et al. Hydrolysis and Maillard reactions during ripening of Iberian ham[J]. Journal of Food Science, 1992, 57(4): 813-815.
[7]Paris M, Jones M G, Eglinton J K. Genotyping single nucleotide polymorphisms for selection of barley β-amylase alleles[J]. Plant molecular biology reporter, 2002, 20(2): 149-159.
[8]夏巧玉, 江均平, 朱利泉, 等. 大豆耐热 β-淀粉酶基因在大肠杆菌中的表达[J]. 西南农业大学学报, 2006, 28(4): 553-557.
[9]Reddy R M, Reddy P G, Seenayya G. Enhanced production of thermostable β-amylase and pullulanase in the presence of surfactants byClostridiumthermosulfurogenesSV2[J]. Process Biochemistry, 1999, 34(1): 87-92.
[10]陈军营, 朱雪萍, 尹海燕, 等. 小麦高分子量谷蛋白亚基 1Dx5 基因在大肠杆菌中的表达[J]. 麦类作物学报, 2007, 27(4): 597-602.
[11]田亚平, 郭鸿飞, 肖光焰, 等. 一种麦芽 β-淀粉酶的纯化和特性研究[J]. 食品工业科技, 2003, 24(9): 22-24.
[12]亚平. 生化分离技术[M].北京:化学工业出版社, 2006.
[13]MATHEWSON P R, Seabourn B W. A new procedure for specific determination of beta.-amylase in cereals[J]. Journal of Agricultural and Food Chemistry, 1983, 31(6): 1 322-1 326.
[14]Mccleary B V, Codd R. Measurement of β-amylase in cereal flours and commercial enzyme preparations[J]. Journal of Cereal Science, 1989, 9(1): 17-33.
[15]靳纪培. 麦芽中 β-淀粉酶的提取, 纯化及应用研究[D].无锡:江南大学, 2009.
[16]YOSHIGI N, OKADA Y, SAHARA H, et al. Expression inEscherichiacoliof cDNA encoding barley β-amylase and properties of recombinant β-amylase[J]. Bioscience, Biotechnology, and Biochemistry, 1994, 58(6): 1 080-1 086.
[17]FONSECA-MALDONADO R, VIEIRA D S, ALPONTI J S, et al. Engineering the pattern of protein glycosylation modulates the thermostability of a GH11 xylanase[J]. Journal of Biological Chemistry, 2013, 288(35): 25 522-25 534.
Heterologous expression of β-amylase gene from barley inE.coli
ZHU Pei1,LI Qi2,ZHU Lin-jiang3,HE Peng-yu4,LI Yong-xian1*
1(Lab of Brewing Science and Technology, Jiangnan University, Wuxi 214122, China)2(School of Biotechnology, Jiangnan University, Wuxi 214122, China)
ABSTRACTBarley β-amylase is widely applied in food and brewing industries due to its high activities, good stability and performance. However, the low production rate and high price of barley β-amylase limit its application. In this study, the barley β-amylase was heterologously expressed in Escherich coli. The β-amylase gene was codon-optimized, inserted into plasmid pET28a (+) and over-expressed in E.coli BL21 (DE3) cells. After comparing the recombinant β-amylase with the native one from barley, the results showed that the molecular weight of the recombinant β-amylase was similar with that of the native barley β-amylase, which was about 59 kDa. Its specific activity was decreased to 285.5U/mg, which was lower than that of the enzyme from barley (588 U/mg). Furthermore, its optimum temperature was declined 10 ℃, while its optimum pH kept consistent. Therefore, barley β-amylase gene could be effectively over-expressed in bacterial cells, while its enzymatic properties altered greatly.
Key wordsbarley; β-amylase; heterologous expression; Escherichia coli
DOI:10.13995/j.cnki.11-1802/ts.201606006
基金项目:国家高技术研究发展计划(863计划);中央高校基本科研业务费资金和江苏高校优势学科建设工程项目
收稿日期:2015-12-09,改回日期:2016-01-20
第一作者:硕士研究生(李永仙为通讯作者,E-mail:yxli@jiangnan.edu.cn)。
