产碳青霉烯酶肺炎克雷伯菌的耐药基因及流行病学研究进展
2015-11-24刘婧娴
刘婧娴, 俞 静, 刘 瑛
·综述·
产碳青霉烯酶肺炎克雷伯菌的耐药基因及流行病学研究进展
刘婧娴, 俞 静, 刘 瑛
肺炎克雷伯菌; 碳青霉烯酶; 耐药基因; 流行病学
肠杆菌科细菌对碳青霉烯类抗生素的耐药机制主要包括产碳青霉烯酶、外膜蛋白缺失或突变、外排泵过度表达以及青霉素结合蛋白变异等,其中最主要的是产碳青霉烯酶。虽然各种肠杆菌科细菌的耐药机制大体相同,但是各种耐药基因在不同种属细菌中的分布和流行情况并不相同。肺炎克雷伯菌是临床上最常见的条件致病菌之一,也是检出率最高的耐碳青霉烯类肠杆菌科细菌。由于临床上治疗耐碳青霉烯类肺炎克雷伯菌感染的药物非常有限,此类患者的病死率较高[1]。自1997年在美国北卡罗莱纳州发现第1株耐碳青霉烯类抗生素的肺炎克雷伯菌以来[2],对碳青霉烯类耐药或敏感性降低的肺炎克雷伯菌检出率呈持续上升趋势,在肺炎克雷伯菌中发现的耐药基因突变类型也越来越多,这些基因大多存在细菌质粒上,往往通过细菌间的传播导致医院感染的流行。
A m bler分类法根据氨基酸序列的相似性将β内酰胺酶分为A、B、C、D 4类,其中A、B、D类中具有水解碳青霉烯类活性的酶被称为碳青霉烯酶。肺炎克雷伯菌对碳青霉烯类抗生素敏感性下降的最主要原因是产碳青霉烯酶。
1 A类碳青霉烯酶
A类碳青霉烯酶的催化基团活性部位均含丝氨酸残基,可以被克拉维酸和他唑巴坦抑制。目前,A类碳青霉烯酶共分为6组:K P C、G ES、S M E、I M I/ N M C-A、S H V-38和 SF C-1[3],各组酶甚至同一组中的不同种酶,对碳青霉烯类的水解活性有很大差异。在肺炎克雷伯菌中发现的A 类酶包括 K P C型、G ES型及S H V-38型等,见表1。
产K P C型碳青霉烯酶是肺炎克雷伯菌对碳青霉烯类抗生素耐药最常见的机制,目前已知的 K P C酶共有14个突变类型(K P C-1和 K P C-2被证实是同一种酶),在肺炎克雷伯菌中已发现其中11种,以 K P C-2和 K P C-3较为常见。K P C-3在西方国家检出率较高,如意大利曾多次出现产 K P C-3酶肺炎克雷伯菌的暴发流行[4]。跨国旅行的患者与携带K PC基因的肺炎克雷伯菌在不同国家之间的传播有密切关系。2012年澳大利亚报道了第1例产K PC酶肺炎克雷伯菌临床分离株,该患者曾去过希腊[5],此后诊断为这类细菌感染的患者也多有其他国家旅行经历[6]。法国 M eaux医疗中心曾出现过产 K PC-3酶肺炎克雷伯菌的 小 规模 流 行,M LST 分型 均 为ST512,最后证实与一名来自意大利的患者有关[7]。我国于2007年发现第1例产K PC-2酶的肺炎克雷伯菌[8],近年浙江地区先后在肺炎克雷伯菌中检测出K PC-12、K PC-2-l ike等 新 的 突变类型[9]。Hawser等[10]回溯性研究2008—2009年世界抗生素耐药性趋势监测(S M A R T)中分离自腹腔感染患者的肺炎克雷伯菌时,发现来自希腊的4株肺炎克雷伯菌均携带有blaKPC-11基因。除此之外 在肺炎 克雷伯菌 中发现的K PC酶亚型,如 K PC-4、5、6、7、8、14等,都仅在美国检出,其他地区尚无流行的报道。
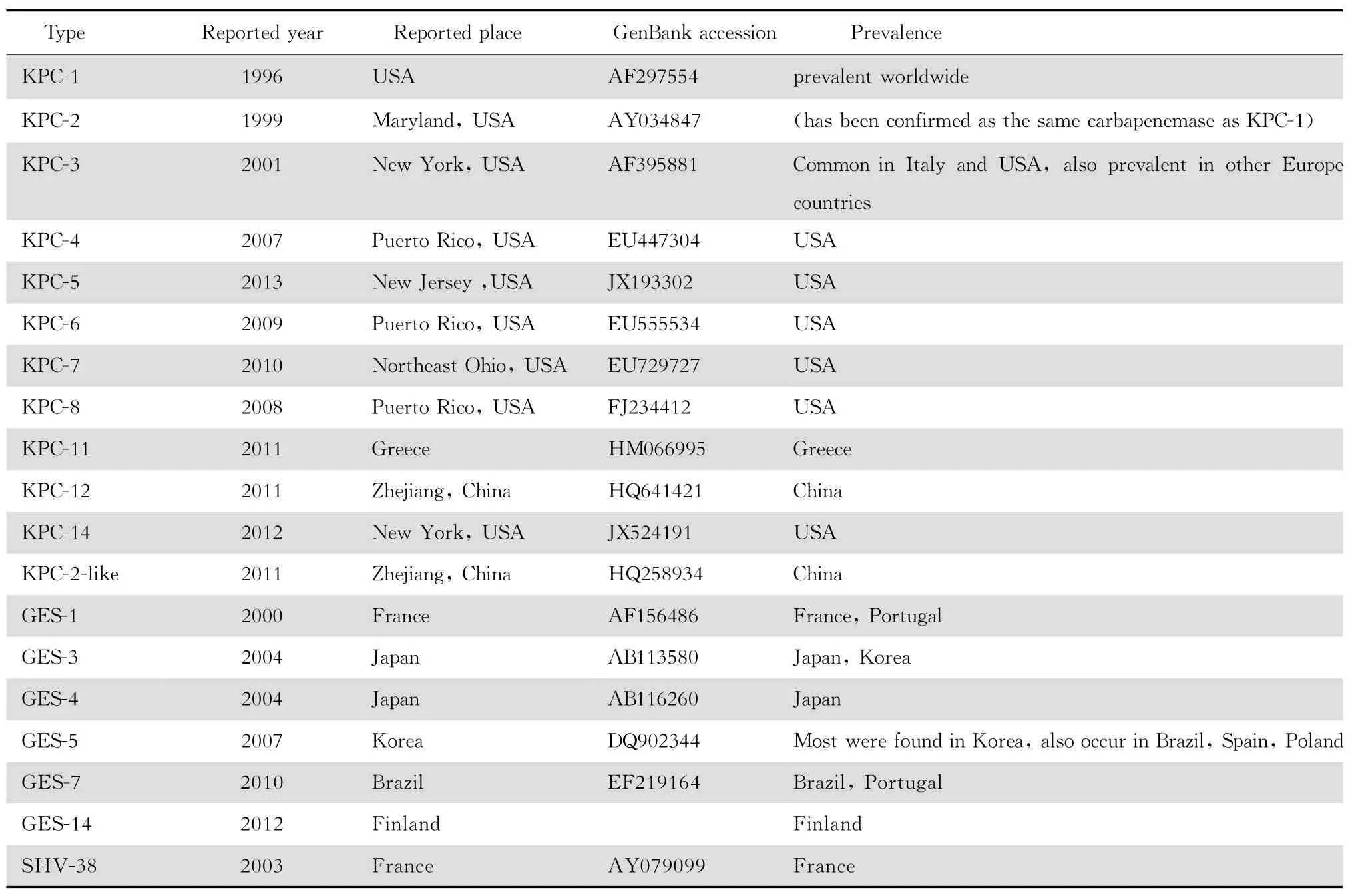
表1 在肺炎克雷伯菌中发现的A类碳青霉烯酶Table 1 Class A carbapenemases identified inKlebsiella pneumoniae
G ES酶最初 被 认为是 超广谱β 内 酰 胺 酶(ESB L),但后来发现其中某些亚类具有低水平水解亚胺培南的能力,所以这些亚类的G ES酶被归入 A类碳青霉烯酶。目前G ES型酶的检出率较低,可能是由于G ES酶对于碳青霉烯类水解能力不强,携带blaGES基因的 临床 分离 株耐 药性 较弱,从而导致漏检。然而G ES酶的分布范围较广,在希腊、法国、葡萄牙、南非、法属圭亚那、巴西、阿根廷、韩国和日本均有报道。肺炎克雷伯菌中与碳青霉烯类耐药相关的G ES酶有G ES-1、G ES-3、G ES-4、G ES-5、G ES-7、G ES-14等类型。其中 G ES-5相对较为多见,该酶的编码基因存在于I类整合子上,对于亚胺培南的水解活性较 G ES-2、G ES-4更强。最初是在韩国分离 到 的1 株 肺炎 克 雷伯 菌 中检 出blaGES-5基因[11],现在巴西、西班牙、波兰等地也开始流行。2000年在法属圭亚那 首 次 发 现 携带blaGES-1基因 的 肺 炎 克 雷伯菌,之后葡萄牙报道了产G ES-1酶肺炎克雷伯菌的医院感染暴发流行[12]。G ES-3、G ES-4仅见于亚洲 地 区,其 中日本 检出较 多[13]。而 携带blaGES-7基 因的肺炎克雷伯菌目前仅巴西和葡萄牙分别报道过1例[14]。芬兰2008—2011年耐药监测网调查中发现了世界范围内唯一1株产G ES-14酶的肺炎克雷伯菌[15]。目前中国尚无关于产G ES酶的肺炎克雷伯菌检出的报道。
2 B类碳青霉烯酶
B类碳青霉烯酶又称金属β内酰胺酶(M B L),所有的金属酶都含有至少1个锌离子活性中心,可以水解包括碳青霉烯类在内带有β内酰胺环的多种抗生素,其水解活性不被克拉维酸、舒巴坦等酶抑制剂抑制,但可以被乙二胺四乙酸(E D T A)及巯基化合物等金属离子螯合剂所抑制。根据基因序列的差异,金属酶可分为3个亚类:B1,B2和B3,目前在肺炎克雷伯菌中检测到的金属酶都属于B1亚类,主要有I M P、VI M和N D M 等类型。见表2。
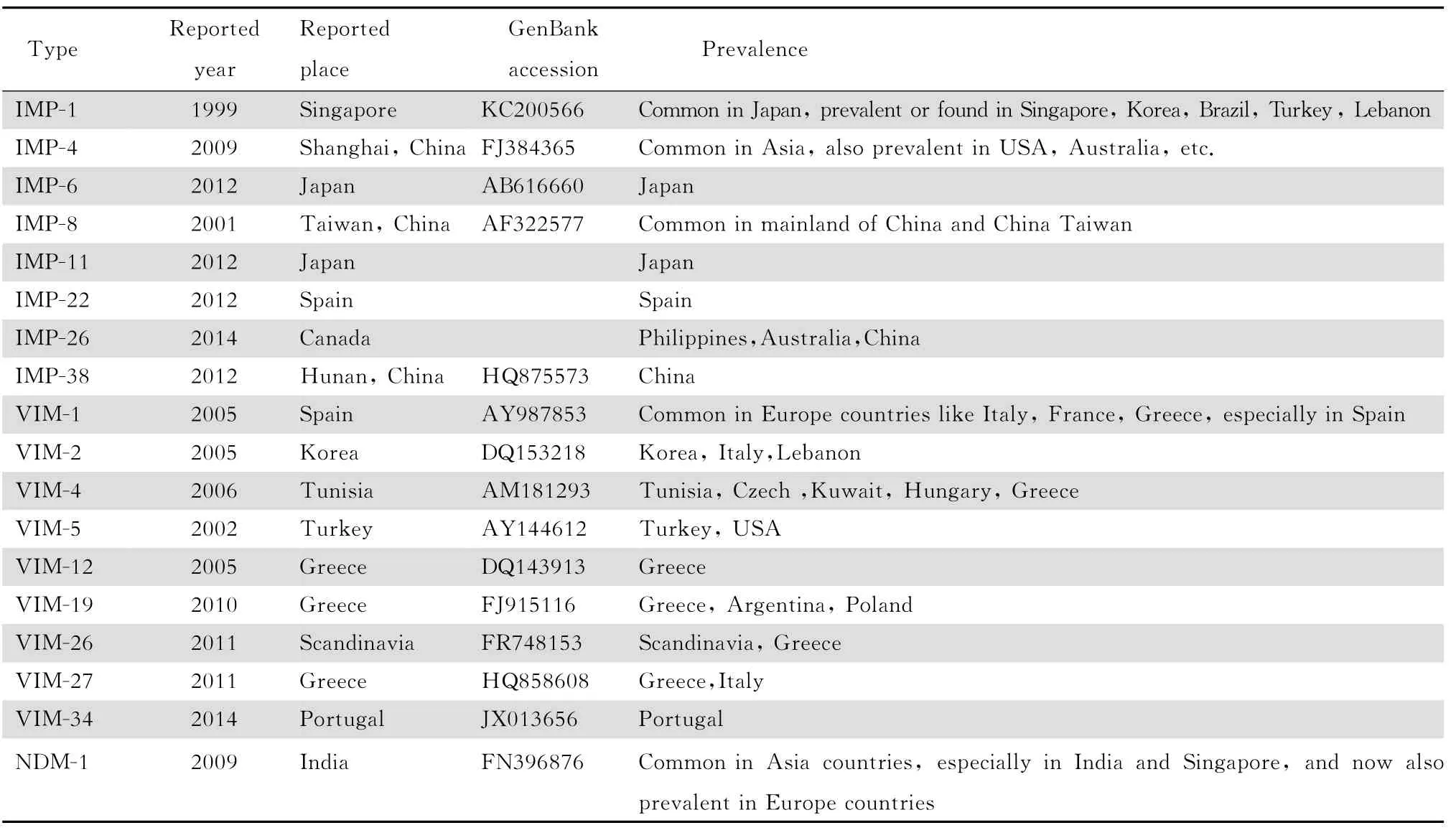
表2 在肺炎克雷伯菌中发现的B类碳青霉烯酶Table 2 Class B carbapenemases identified inKlebsiella pneu moniae
迄今为止已发现的I M P酶共有38个类型,该家族大多由I M P-1、I M P-2突变而来,同源性较高,主要通过I类整合子进行水平传播。I M P酶本身对碳青霉烯类抗生素水解能力并不强,携带blaIMP基因的细菌并不一定在药敏试验中显示对碳青霉烯类耐药。产I M P酶肺炎克雷伯菌临床分离株呈现出对碳青霉烯类高度耐药,往往合并有外膜蛋白缺失等其他耐药 机制[16]。在肺 炎克 雷 伯菌中 检 出的I M P类型有I M P1、4、6、8、11、22、26、32和38等。最初,产I M P酶的肺炎克雷伯菌仅见于日本及一些东南亚国家,现在包括中国在内的整个东南亚,以及澳大利亚、美国、加拿大、巴西、英国、意大利、葡萄牙等国家均有检出。产I M P-1酶肺炎克雷伯菌曾多次在日本引起医院感染暴发流行[17],此外,韩国、新加坡、土耳其、巴西等也有检出或流行的报道。我国较为常见的类型是I M P-4、I M P-8,其中浙江地区分离率最高[18]。湖南曾有产I M P-38酶肺炎克雷伯菌的小规模流行[19],此类菌株几乎都携带有多种毒力基因,具有黏附性强、易在呼吸道和医疗器械定植的特点,并且能产生肠毒素和耶尔森毒素,致病力很强[20]。产I M P-26酶肺炎克雷伯菌在中国、菲律宾及澳大利亚均有检出,但是不同国家间流行的菌株并 不 具有 同 源性[21]。
VI M 是所有类型金属酶中对碳青霉烯类的水解能力最强的一种,最初VI M 型金属酶在铜绿假单胞菌中流行最为广泛,但是自2003年希腊报道了第1例产VI M-1金属酶的肺炎克雷伯菌以来[22],其他类型的VI M 也相继在肺炎克雷伯菌中被检出。目前报道的 VI M 突变类型共有34种,在肺炎克雷伯菌中发现的有VI M-1、2、4、5、12、26、27和34等亚类,见表2,其中VI M-1最为常见。虽然表达 VI M-1酶的菌株对于青霉素类、头孢菌素类、碳青霉烯类的敏感性均显著下降,却对氨曲南保持敏感性。VI M 基因通常借助I类整合子在不同种属细菌间快速传播,流行范围广,并有明显的地区特性。欧洲绝大部分国家均有携带blaVIM基因的肺炎克雷伯菌的检出甚至流行,尤其是西班牙、希腊和意大利[23-24]。 除欧 洲 外,在韩 国也 曾 检出 过 1 株 产VI M-2酶肺炎克雷伯菌,M LS T分型为S T70,与韩国流行的肺炎克雷伯菌类型亲缘关系甚远,可能是由其他地区传入[25]。产 VI M-4酶肺炎克雷伯菌的检出率虽然不高,但是其流行范围涵盖了欧洲、亚洲、非洲国家,如突尼斯、科威特、匈牙利等。VI M-12是一种较为特殊的碳青霉烯酶,虽然其氨基酸序列与 VI M-1的 N端及VI M-2的C端高度相似,但对水解底物具有特异性,仅对青霉素类及亚胺培南具有水解活性[26]。有关产 VI M-12酶肺炎克雷伯菌的报道并不多,主要集中在希腊,曾在希腊的一所教 学 医院 引 起流 行[27]。
2009年,Yong等[28]报道了从肺炎克雷伯菌中首次分离出新的金属酶基因型即 N D M-1,该菌株对几乎所有β内酰胺类耐药,仅对替加环素和多黏菌素敏感。携带blaNDM-1基因 的 肺 炎克 雷 伯菌 在 亚洲地区较为多见,目前在印度、摩洛哥、中国大陆、中国台湾、新加坡、韩国、日本、科威特、阿曼苏丹国等均有报道。BlaNDM-1基因 在欧洲 和 北美 洲 传播 和 流行也与患者的“跨国医疗旅游”有关。Y ong等[28]报道的瑞典患者是在印度旅行过程中感染了产N D M-1的肺炎克雷伯菌,而后法国报道的1例感染产N D M-1金属酶的肺炎克雷伯菌的患者则来自伊拉克,美国报道的blaNDM-1携带株多分离自印度或 巴勒斯坦的患者[29]。这也表明印度及中东地区很有可能是 N D M-1的起源及聚集地,最近也有报道称西欧地区N D M-1 的流行可能来 自南欧巴尔干地区[30]。携带 N D M-1基因的肺炎克雷伯菌在我国尚不多见,然而检出率呈逐年增多趋势,现在江西、北京、海 南、湖 南 等 地 均有报道[31-33],虽然暂未 出 现 大规模流行,但仍需警惕。
3 D类碳青霉烯酶
D类碳青霉烯酶又称苯唑西林酶(O X A酶),大多对碳青霉烯类水解效率较低,不能水解氨曲南,常与其他耐药机制协同导致细菌对碳青霉烯类高度耐药。目前发现的 O X A酶有200多种,在肺炎克雷伯菌中检出最多的为O X A-48,其次是O X A-181,其他类型如 O X A-1、O X A-30、O X A-41等也与肺炎克雷伯菌对碳青霉烯类耐药有关。见表3。O X A-48与O X A-181酶的编码基因均位于质粒上,两者仅有4个氨基酸的差异,抗生素水解谱相似。起先关于产O X A-48酶的肺炎克雷伯菌的检出及医院感染暴发流行的报道几乎都集中于土耳其[34],现已在埃及、英国、突尼斯、法国、以色列、西班牙、新西兰、利比亚等地传播甚至引起暴发流行。产O X A-181酶肺炎克雷伯菌的报道则多来自新加坡、澳大利亚、新西兰、罗马尼亚、南非、印度等地。O X A-41是第1个在肺炎克雷伯菌中发现的O X A类酶,仅在法国有过1例报道,与其他类型不同的是O X A-41酶可以高效催化亚胺培南水解。产O X A-247酶肺炎克雷伯菌分离自1例阿根廷患者,此前,该患者曾感染过产O X A-163酶肺炎克雷伯菌,流行病学、生物化学及分子生物学等相关实验证明 O X A-247基因是由O X A-163基因突变而来。其他在肺炎克雷伯菌中发现的O X A 酶,如O X A-41、47、48、72、162、163、244和245等,多在欧洲国家被检出。虽然大部分O X A酶的亚类仅在局部地区检出或小规模的流行,但近年 O X A酶新突变类型出现的速度非常之快,需引起足够的重视。
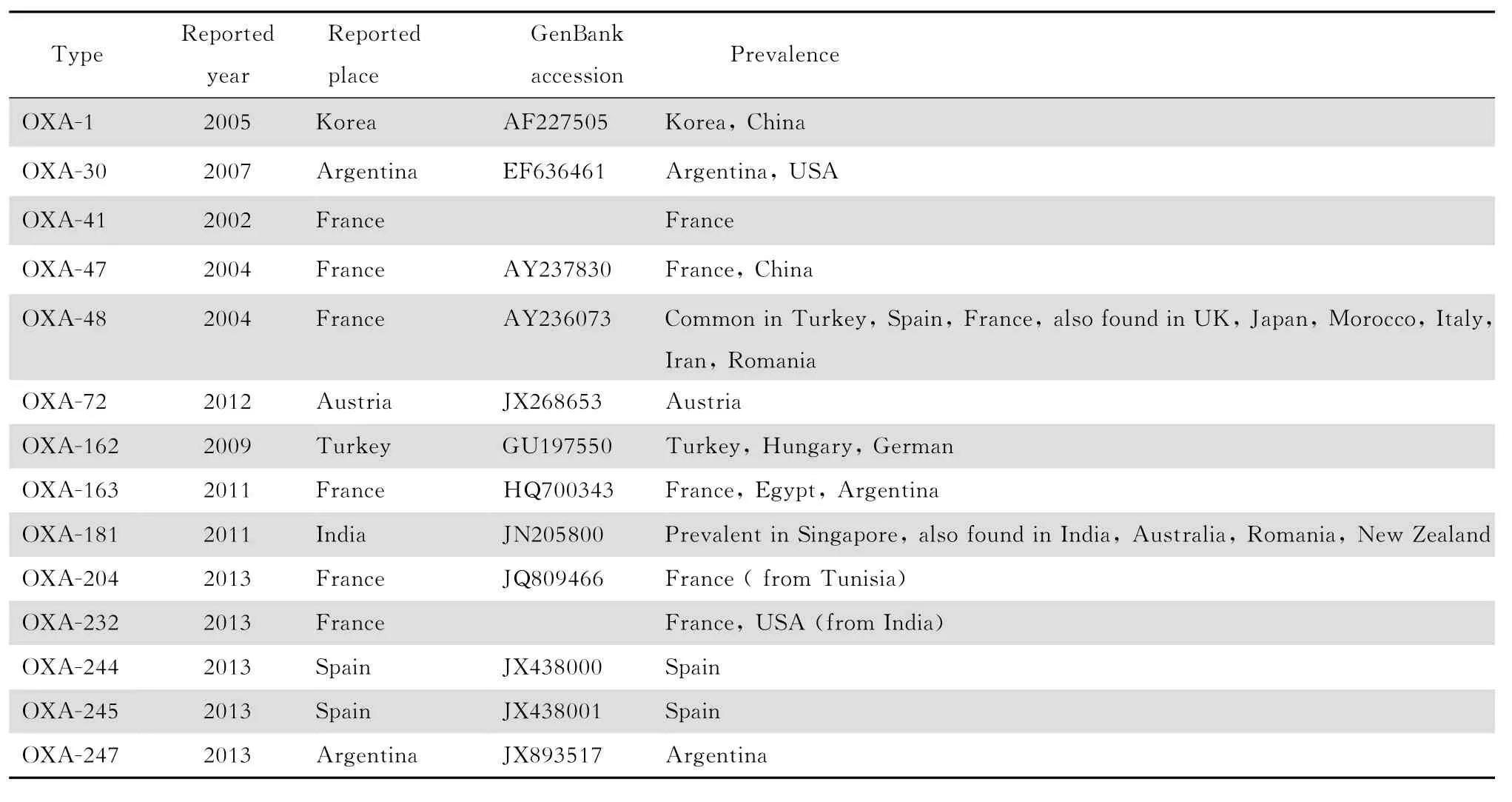
表3 在肺炎克雷伯菌中发现的D类碳青霉烯酶Table 3 Class D carbapenemases identified inKlebsiella pneu moniae
4 总结与展望
肺炎克雷伯菌中检测到的碳青霉烯酶基因种类繁多,每个地区流行的耐药株也各有特点。A类酶中 K PC-2是最常见的耐药基因类型,世界范围内均有流行。除此之外,K PC-3及G ES-5在肺炎克雷伯菌中也分布较广,K PC-3主要见于美国、意大利等西方国家,而 G ES-5则在韩国等亚洲国家较为多见。就B类金属酶而言,目前在肺炎克雷伯菌中检出较多 的 类型 有I M P-1、I M P-4、I M P-8、VI M-1、N D M-1等。金属酶的流行具有较明显的地域性,产I M P-4酶的肺炎克雷伯菌在中国等亚洲地区流行广泛,VI M-1则是希腊等欧洲国家最常见的耐药基因,N D M-1在印度及中东地区较为多见,但是在其他亚洲地区的检出也逐渐增多,同时在欧洲、北美洲等地也呈扩散趋势。D类O X A 酶中,O X A-48和O X A-181较常见。最初 O X A-48酶仅见于土耳其,近年迅速向周边国家扩散,在欧亚地区流行已较为广泛。产 O X A-181酶肺炎克雷伯菌则多分离自新加坡,但是其他国家也相继有检出或流行。虽然大部分在产碳青霉烯酶肺炎克雷伯菌中检出的耐药基因类型影响范围较为局限,但是其检出区域都呈扩大趋势,而且近年有很多新的突变类型相继出现,值得引起广泛关注。
产碳青霉烯酶肺炎克雷伯菌给临床治疗带来了极大的困扰,是引起医院感染患者高病死率的重要因素,一方面是由于有效治疗药物有限,再者治疗过程中这类细菌可能迅速出现对庆大霉素、多黏菌素等抗生素的继发性耐药(secondary resistance)[35]。有报道指出,通过隔离感染患者、改变医务人员的卫生习惯、深度清洁病房和其他医疗设施以及合理应用抗生素等措施,可以有效地控制产碳青霉烯酶肺炎克雷伯菌医院感染暴发流行[36-37]。目前产碳青霉烯酶的肺炎克雷伯菌在世界范围内流行广泛,相应的耐药基因涉及面广,给临床及研究带来很大的挑战,应引起足够的重视。在寻找和开发有效治疗药物的同时,应制定相应的制度以预防医院感染的出现和流行,一旦感染暴发后能及时控制避免进一步扩散。
[1] N ord mann P,Cuzon G,Naas T,et al.T he real threat of Klebsiellapneu moniae carbapenemase-producing bacteria[J].Lancet Infect Dis,2009,9(4):228-236.
[2] Yigit H,Queenan A M,Anderson GJ,et al.Novel carbapenemhydrolyzing beta-lacta mase,K P C-1,fro m a carbapenemresistant strain ofKlebsiella pneu moniae[J].A ntimicrob A gents Chem other,2001,45(4):1151-1161.
[3] W alther-Rasm ussen J,H oiby N.Class A carbapenemases[J].J A ntimicrob Chem other,2007,60(3):470-482.
[4] Pulcrano G,Lula D V,de Luca C,et al.Clonal dissemination of Klebsiellapneu moniae S T512 carrying blaK P C-3in a hospitalin southern Italy[J].A P MIS,2014,122(1):42-46.
[5] Coatsworth N R,H untington P G,H ardiman R P,et al.A case of carbapenemase-producingKlebsiella pneu moniaein A ustralia[J].Pathology,2012,44(1):42-44.
[6] Chua K Y,Grayson M L,Burgess A N,et al.T he growing burden of m ultidrug-resistant infections a m ong returned A ustralian travellers[J].M ed J A ust,2014,200(2):116-118.
[7] Cuzon G,Naas T,Demachy M C,et al.N osoco mial outbreak ofKlebsiellapneu moniae harbouringblaKPC-3in France subsequent to a patient transfer fro m Italy[J].J A ntimicrob A gents,2012,39(5):448-449.
[8] W ei Z Q,Du X X,Y u Y S,et al.Plasmid-mediated K P C-2 in a Klebsiella pneu moniaeisolate fro mChina[J].A ntimicrob A gents Chem other,2007,51(2):763-765.
[9] Zhu J M,Jiang RJ,Mi Z H,et al.N ovel K PC variant fro m a carbapenem-resistant strain ofKlebsiella pneu moniaein a Chinese hospital[J].Infect Control H osp Epidemiol,2011,32(10):1050-1052.
[10] H awser SP,Bouchillon S K,Lascols C,et al.Susceptibility ofKlebsiellapneu moniae isolatesfro m intra-abdo minal infections and m olecular characterization of ertapenemresistant isolates[J].A ntimicrob A gents Chem other,2011,55(8):3917-3921.
[11] Bae IK,Lee Y N,Jeong S H,et al.Genetic and biochemical characterization of G ES-5,an extended-spectru m class A beta-lacta mase fro m Klebsiella pneu monia [J].Diagn Microbiol Infect Dis,2007,58(4):465-468.
[12] Duarte A,Boavida F,Grosso F,et al.O utbreak of G ES-1 beta-lacta mase-producing m ultidrug-resistant Klebsiella pneu moniaein a university hospitalin Lisbon,Portugal[J]. A ntimicrob Agents Chem other,2003,47(4):1481-1482.
[13] Ryoo N H,Kim E C,H ong S G,et al.Dissemination of S H V-12 and C T X-M-type extended-spectru m beta-lacta mases a m ong clinical isolates ofEscherichia coliandKlebsiella pneu moniaeandemergenceof G ES-3 in K orea[J].J A ntimicrob Chem other,2005,56(4):698-702.
[14] Manageiro V,Ferreira E,Cougnoux A,et al.Characterization of the inhibitor-resistant S H V beta-lacta mase S H V-107 in a clinicalKlebsiellapneu moniae straincoproducingG ES-7 enzy me[J].A ntimicrob A gents Chem other,2012,56(2):1042-1046.
[15] Österblad M,Kirveskari J,Hakanen AJ,et al.CarbapenemaseproducingEnterobacteriaceaeinFinland:the first years(2008-11)[J].J A ntimicrob Chem other,2012,67(12):2860-2864.
[16] Sho T,M uratani T,H amasuna R,et al.T he mechanism of high-level carbapenem resistance inKlebsiella pneu moniae:underlying o m pk36-deficient dtrains represent a threat of emerginghigh-level carbapenem-resistantK.Pneu moniae with I M P-1 beta-lacta mase production in Japan[J].Microb Drug Resist,2013,19(4):274-281.
[17] Sugimoto T,Kimura S,Nakama E,et al.Increase of metal lobeta-lactamase producing bacteria positive cases including Klebsiella pneu moniaein a Japanese university hospital[J]. Rinsho Byori,2010,58(6):553-558.
[18] Yang Q,W ang H,Sun H,et al.Phenotypic and genotypic characterization of Enterobacteriaceae with decreased susceptibility to carbapenems:results fro m large hospitalbased surveillance studies in China[J].A ntimicrob A gents Chem other,2010,54(1):573-577.
[19] Liu W.A novel I M P metallo-beta-lacta mase,I M P-38,fro m an outbreak of aK.pneu moniaclone strain in a university hospitalin central China[C].52nd Interscience Conference on A ntimicrobial A gents and Chem otherapy.San Francisco,C A,U S A:A mer Soc Microbiol,2012:C2083.
[20] 谷秀梅,刘文恩,简子娟,等,携带产新型碳青霉烯酶基因blaIM P-38肺炎克雷伯菌致病性分析[J].临床检验杂志,2013,31(9):698-705.
[21] Peirano G,Lascols C,H ackel M.M olecular epidemiology of Enterobacteriaceaethat produce VI M s and I M Ps fro m the S M A R T surveillance program[J].Diagn Microbiol Infect Dis,2014,78(3):277-281.
[22] Giakkoupi P,Xanthaki A,Kanelopoulou M,et al.VI M-1 M etallo-beta-lacta mase-producingKlebsiella pneu moniae strains in Greek hospitals[J].J Clin Microbiol,2003,41(8):3893-3896.
[23] Pena I,Picazo JJ,Rodríguez-Avial C,et al.CarbapenemaseproducingEnterobacteriaceaein a tertiary hospitalin M adrid,Spain:high percentage of colistin resistance am ong VI M-1-producingKlebsiella pneu moniaeS T11 isolates[J].Int J A ntimicrob A gents,2014,43(5):460-464.
[24] Oteo J,Saez D,Bautista V,et al.Carbapenemase-producing Enterobacteriaceaein Spain in 2012[J].A ntimicrob A gents Chem other,2013,57(12):6344-6347.
[25] Kim SY,Shin J,Shin SY,et al.Characteristics of carbapenemresistantEnterobacteriaceaeisolates fro mK orea[J].Diagn Micr Infec Dis,2013,76(4):486-490.
[26] Kontou M,Pournaras S,Kristo I,et al.M olecular cloning and biochemical characterization of VI M-12,a novel hybrid VI M-1/VI M-2 metallo-beta-lacta masefro m a Klebsiella pneu moniae clinical isolate, reveal atypical substrate specificity[J].Biochemistry,2007,46(45):13170-13178.
[27] Tokatidou D,Tsivitamidou M,Pournaras S,et al.O utbreak caused by a m ultidrug-resistantKlebsiella pneu moniaeclone carrying blaVI M-12 in a university hospital[J].J Clin Microbiol,2008,46(3):1005-1008.
[28] Yong D,Toleman M A,Giske C G,et al.Characterization of a new metallo-beta-lacta mase gene,bla(N D M-1),and a novel erythro m ycin esterase gene carried on a unique genetic structure inKlebsiella pneu moniaesequence type 14 fro m India[J].A ntimicrob Agents Chem other,2009,53(12):5046-5054.
[29] M ochon A B,Garner O B,Hindler JA,et al.New Delhi metallo-β-lacta mase (N D M-1 )-producing Klebsiella pneu moniae:case report and laboratory detection strategies[J].J Clin Microbiol,2011,49(4):1667-1670.
[30] M azzariol A,Bošnjak Z,Bal larini P,et al.N D M-1-producing Klebsiella pneu moniae,Croatia[J].E merg Infect Dis,2012,18(3):532-534.
[31] Zhou G,G uo S,Luo Y,et al.N D M-1-producing strains,fa milyEnterobacteriaceae,in hospital,Beij ing,China[J]. E merg Infect Dis,2014,20(2):340-342.
[32] 李天娇,王旭明,符生苗,等.海南发现4株产 N D M-1多重耐药的肺炎克雷伯菌[J].中国热带医学,2013,13(12):1468-1474.
[33] 邹明祥,邬靖敏,李军,等,产 N D M-1肺炎克雷伯菌中国分离株的初步研究[J].中国当代儿科杂志,2012,14(8):616-621.
[34] Carrër A,Poirel L,Eraksoy H,et al.Spread of O X A-48-positive carbapenem-resistantKlebsiella pneu moniaeisolates in Istanbul,Turkey[J].A ntimicrob Agents Chem other,2008,52(8):2950-2954.
[35] Lübbert C,Faucheux S,Becker-Rux D,et al.Rapid emergence of secondary resistance to genta micin and colistin following selective digestive deconta mination in patients with K P C-2-producingKlebsiella pneu moniae:a single-centre experience[J].Int J A ntimicrob Agents,2013,42(6):565-570.
[36] Gaibani P,Colombo R,Arghittu M,et al.Successful containment and infection control of a carbapenem-resistantKlebsiella pneu moniae outbreakin anItalian hospital [J].New Microbiol,2014,37(1):87-90.
[37] T ho mas CP,M oore LS,Ela min N,et al.Early(2008-2010)hospital outbreak ofKlebsiella pneu moniaeproducing O X A-48 carbapenemase in the U K[J].Int J A ntimicrob A gents,2013,42(6):531-536.
Research update on the resistant genes and epidemiology of carbapenemaseproducingKlebsiella pneumoniae
LIU Jingxian,Y UJing,LIUYing. (Department ofLaboratoryM edicine,XinhuaH ospital,Shanghai Jiao Tong University School of M edicine,Shanghai 200092,China)
R378
A
1009-7708(2015)01-0091-06
2014-05-28
上海市卫生局资助课题(2010024)。
上海交通大学医学院附属新华医院检验科微生物室,上海 200092。
刘婧娴(1988—)女,硕士在读,主要从事肠杆菌科细菌对碳青霉烯类抗生素耐药机制研究。
刘瑛,E-mail:liuying0129@g mail.co m。
