运动性骨骼肌损伤后不同时刻大鼠肱三头肌超微结构及肌酸激酶的变化
2015-06-06孙良,赵刚
孙 良,赵 刚
(吉林师范大学,吉林四平136000)
运动性骨骼肌损伤后不同时刻大鼠肱三头肌超微结构及肌酸激酶的变化
孙 良,赵 刚
(吉林师范大学,吉林四平136000)
通过建立大鼠下坡跑运动损伤模型,研究运动损伤后骨骼肌超微损伤和恢复发生的机制。主要结果:运动后大鼠肱三头肌超微结构发生改变,电镜下可见肌纤维排列紊乱,Z线异常(包括Z线流、Z线模糊、Z线扭曲和Z线消失),肌细胞膜模糊、溶解,线粒体肿胀、空泡样改变等。这些现象随运动时间的不同而发生改变。运动后24 h表现最为明显,而后逐渐恢复;运动后72 h恢复明显,但没有完全恢复。运动后即刻血清CK活性(2035.42±426.49)U/L与对照组血清CK活性(293.66±76.07)U/L相比,非常显著增高(P<0.01),达到了峰值,而后逐渐恢复。运动后72h组血清CK活性(425.51±143.34)U/L与即刻组血清CK活性相比已显著恢复(P<0.01)。结论:运动后24 h骨骼肌的损伤程度最为严重,72 h后可明显恢复;大强度运动对CK的活性具有非常显著的影响,血清CK活性的变化能够反映运动后骨骼肌超微结构损伤的状况。
运动骨骼肌损伤;骨骼肌超微结;肌酸激酶
运动性骨骼肌损伤(EIMI)发生机制主要是训练或运动时,由于局部肌肉负荷过大或过度用力,超过机体的承受能力,运动后肌肉功能未得到充分恢复,以及运动与恢复发生失衡等原因所致。EIMI是运动训练和体育锻炼中常见的现象,主要表现为肌紧张、放松能力降低、伴有疼痛肿胀感觉。EIMI经常伴随着一系列血清酶、肌纤维超微结构改变,主要表现为肌节缩短、肌丝排列改变、粗肌丝相互位置紊乱、部分肌丝断裂或消失,以及Z带扭曲、增宽、部分或全部消失等现象。大量研究发现,EIMI出现后,肌酸激酶(creatine kinase,CK)活性随之发生敏感的变化。有学者认为[1]测定血清CK活性对监测运动员疲劳的出现和恢复程度具有实践意义。本研究通过大鼠跑台运动损伤模型,研究EIMI过程中的骨骼肌超微结构变化,以及CK在损伤中表达的变化规律,力图为制定科学合理的运动训练和健身方案提供依据。
1 研究对象与方法
1.1 研究对象
选取雄性SD健康大鼠48只,平均体重(298.13g±12.68)g(由某医学院实验动物中心提供)。筛选具有正常跑台下坡能力的大鼠进行分组,随机分为安静对照组(C)、运动后即刻组(E0)、运动后12 h组(E1)、运动后24 h组(E3)、运动后48 h组(E3)和运动后72 h组(E4)6组,每组8只大鼠。各组实验大鼠体重无显著性差异,分笼饲养,自由饮食,饲料由某医学院实验动物中心提供。室温(22± 3)℃,湿度55%±5%,光照时间:7:00~19:00。所有动物在实验前均未进行过跑台运动。
1.2 研究方法
1.2.1 力竭模型
实验进行前3天,对受试大鼠进行适应性训练,每天进行5~10 min的跑台练习,速度控制在5~10 m/min,跑台坡度为0°。实验开始时,所有运动组大鼠均在专用电动跑台上进行持续性下坡跑运动,速度为16 m/min,坡度为-16°。运动100 min后休息5 min,然后再运动100 min,总运动时间为200 min。运动组大鼠开始状态都在跑道前端运动,步态正常,跑速均匀。随着运动时间的增加,跑步姿势逐渐发生变化,表现为四肢无力、跑速不均匀、腹部与跑道面接触,甚至出现卧位跑,逐渐向跑道后端移动。运动到近200 min时,大鼠反应明显迟缓,腹部皮毛湿润,四肢足部肿胀;所有大鼠都在跑道后端,跑速明显不均匀,不能保持皮带的速度。运动结束后,大鼠表现为神态倦怠,呼吸困难,反应迟钝,几乎没有逃避反应。根据大鼠力竭的标准[2],可以判断大鼠已经力竭。
1.2.2 动物标本取材及血样检验
标本的选取时间分别在运动后即刻、12 h、24 h、48 h、72 h。大鼠在处死前先称取体重,通过腹腔注射戊巴比妥溶液麻醉,麻醉剂量为0.25~0.5 g/Kg体重。每组取8只大鼠,麻醉后迅速取右侧前肢肱三头肌内侧头中下1/3处约0.5×0.5×1 cm3肌组织块,迅速放入2.5%戊二醛固定液中固定,以备制作透射电镜切片。每组取8只大鼠通过腹主动脉抽取血样,在高速冷冻离心机内经3000转/min离心10 min,取血清于-80℃低温冰箱储存,直到检测。
1.2.3 透射电子显微镜标本制作及观察
1.2.3.1 标本制作步骤 1)骨骼肌标本2.5%戊二醛固定;2)0.1MPBS(磷酸缓冲液,PH为7.2~7.4)漂洗3次,10分/次,4℃;3)1%锇酸(1%OsO4)后固定1.5 h,4℃;4)0.1MPBS(磷酸缓冲液,PH为7.2~7.4)漂洗3次,10 min/次,4℃;5)50%酒精-70%酒精-80%丙酮-90%丙酮-100%丙酮梯度脱水;6)Epon812(环氧树脂812)浸透1夜(室温),然后在35℃~45℃~60℃温度下包埋聚合,各24 h;7)LKB超薄切片机超薄切片(70 nm);8)醋酸铀和柠檬酸铅染色各30分;9)37℃温箱干燥。
1.2.3.2 标本观察与记录每组随机选取8只大鼠的肱三头肌制备电子显微镜切片。每一个电子显微镜切片首先在放大2 000倍的低倍视野内确定观察区域,然后在放大8 000~10 000倍的视野内观察肌纤维的损伤情况。做好观察记录,选好范围拍片,准确记录底片号码,在电脑中备份。
1.2.4 血清肌酸激酶(CK)的测定
CK活性测定采用南京建成生物工程研究所提供的CK试剂盒和6010紫外可见分光光度计进行测定。
测定原理:CK催化三磷酸腺苷和肌酸生成磷酸肌酸,后者很快全部水解为磷酸。此时三磷酸腺苷和二磷酸腺苷仍稳定,加入钼酸铵可形成磷钼酸,进一步可还原成钼蓝,根据生成无机磷的量可推算出酶的活性。
测定方法:取血清20μl,严格按照试剂盒说明书所示程序进行操作,经反应后在660 nm处1 cm光径比色,用蒸馏水调零,测定标准管、标准空白管、测定管和测定空白管的吸光度。按以下公式计算血清CK活性。标准品的浓度为0.05 mg/ml。lmg的酶活力为35活力单位(U)。
CK活性(U/L)=标准品的酶活性(0.05×35 U/L)×(测定管OD值-测定空白管OD值)/(标准管OD值-标准空白管OD值)。
1.2.5 实验数据统计与分析
所有实验数据均采用SPSS 13.0统计分析软件进行处理,测试结果以“平均数±标准差”(珡X±S)表示,采用单因素方差分析和非参数检验进行各组间的差异显著性检验。
2 研究结果
2.1 骨骼肌超微结构变化
2.1.1 安静对照组的骨骼肌超微结构
安静对照组在纵切片上可见肌节结构完整,肌纤维无肿胀,形态正常,肌原纤维排列紧密,A带、H带、I带和M线清晰可见,结构完整,呈明暗相间的横纹。Z线清晰、完整,排列规则。三连管结构正常。肌细胞核膜完整、光滑,核仁清晰,肌核大小正常。肌间线粒体和胞质内线粒体数量较多,大小正常,外膜和嵴完整(图1)。

图1 正常肌小节结构(×12 Kb)
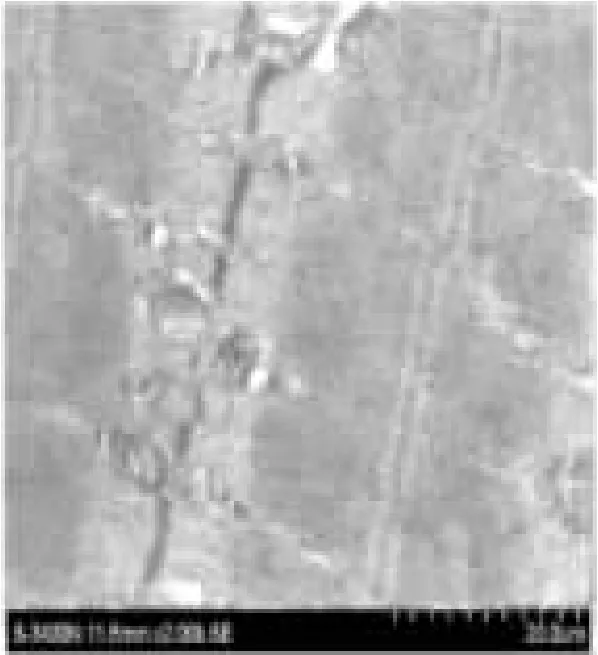
图2 偶见Z线异常(×12 Kb)
2.1.2 运动后即刻组的骨骼肌超微结构变化
运动后即刻组与安静组相比,超微结构发生了一定程度的改变。可见肌纤维排列稍有紊乱,肌丝肿胀。偶见Z线异常。三连管轻度肿胀。肌细胞膜部分模糊、溶解,核膜皱缩、稍肿胀。肌纤维间线粒体肿胀、破裂,偶见空泡样改变,嵴小部分缺损。卫星细胞核皱缩(图2)。
2.1.3 运动后12 h组的骨骼肌超微结构变化
运动后12 h组与运动后即刻组相比,超微结构进一步损伤。可见肌纤维排列紊乱、扭曲,肌纤维间隙增宽,明暗带模糊。Z线局部紊乱、扭曲。横小管肿胀。细胞膜有改变,局部溶解,肌细胞核肿胀,固缩,形状不规则。线粒体空泡变性,部分嵴缺损(图3)。

图3 线粒体空泡变性(×8 Kb)

图4 Z线呈锯齿状(×8 Kb)
2.1.4 运动后24 h组的骨骼肌超微结构变化
运动后24h组超微结构的损伤程度更加明显。可见肌纤维排列不规则、紊乱、扭曲,肌丝间肿胀,明暗带模糊,无法区分A、I带。Z线排列不规则,可见Z线流,Z线呈锯齿状,Z线局部消失。肌细胞膜固缩、部分溶解,肌细胞核浓缩、核膜模糊。线粒体多见空泡变性、肿胀、嵴部分缺损、紊乱(图4)。
2.1.5 运动后48 h组的骨骼肌超微结构变化
运动后48 h组比运动后24 h组超微结构的损伤程度有所减轻。可见肌纤维间肌丝水肿。Z线局部错位,可见Z线流变。肌细胞膜溶解、皱缩,核膜肿胀、模糊。线粒体絮状物改变,外膜局部破坏,嵴部分缺损,常见空泡变性(图5)。
2.1.6 运动后72 h组骨骼肌的超微结构变化
运动后72h组大鼠骨骼肌超微结构损伤程度明显减轻。可见肌纤维排列接近正常,未见肌纤维水肿。肌细胞膜部分皱缩,能够分清明暗带和M线。多数线粒体比较完整,部分线粒体嵴缺损、空泡变性,絮状改变逐渐变化到髓样改变(图6)。
2.2 不同时刻肌酸激酶的变化
实验结果表明,一次性离心力竭运动后即刻血清CK活性比安静时升高6.93倍(P<0.01)。运动后12 h组血清CK活性比安静时升高3.71倍(P<0.01),运动后24 h组血清CK活性比安静时升高2.33倍(P<0.01),运动后48 h组血清CK活性比安静时升高2.06倍(P<0.01),运动后72 h组血清CK活性比安静时升高1.45倍(P>0.05),血清CK活性的变化表明,在运动后即刻血清CK活性达高峰,然后下降,呈恢复的趋势(表1)。

图5 线粒体絮状物改变(×6 Kb)

图6 肌纤维排列接近正常(×10 Kb)

表1 大鼠力竭运动前后血清CK活性的变化(n=8)
3 分析与讨论
上世纪80年代,EIMI引发了体育科研工作者的关注。研究者发现,大负荷运动后骨骼肌产生超微结构变化主要表现为肌节缩短、肌丝排列混乱、Z扭曲模糊等现象。后继研究者对EIMI的过程进行了深入细致的研究。田野等人研究发现,大鼠骨骼肌超微结构损伤在24 h~48 h最为明显,而后逐渐恢复,运动后第7 d时损伤已明显恢复。李世成等人研究发现,实验后骨骼肌超微形态结构在0 h、12 h、24 h和48 h发生了程度不同的改变,其中运动后12~24 h左右最为严重。从本研究结果看,运动后即刻骨骼肌超微结构发生明显变化,出现肌纤维排列紊乱、明暗带模糊、Z线异常等现象。随着运动时间的延长,损伤程度加深,在运动后24 h损伤最为严重,运动后48 h逐渐得以缓解,运动后72 h接近恢复到正常水平。与上述研究基本一致。
CK是一种重要的代谢酶,可通过催化ATP与肌酸可逆反应,参与细胞供能[2]。在正常状态下,CK对肌细胞膜的通透性低,血清CK活性维持在正常范围。大负荷运动后,由于肌细胞膜通透性增加和结构完整性的受损,CK进入血清浓度增加。有学者认为,无论运动强度高低,都会使血清CK活性增加。有研究进一步指出,肌肉收缩方式、运动类型与供能方式不同,导致血清CK的浓度不同。研究认为,抵抗性运动比有氧运动血清中CK的浓度高。无氧运动较有氧运动血清中CK的浓度高。大强度运动较低强度血清中CK的浓度高。但进行马拉松和超长距离跑等运动后,血清CK出现非常显著升高。通常认为,造成这种现象的原因与肌纤维的损伤程度有关。
Takekura[47]等进行大鼠下坡跑实验发现,运动后即刻的CK活性是安静值的3.4倍。运动后4 h CK活性急剧地降低到安静时的水平,但是在运动后16~48 h有个显著的第二次升高(P<0.01)。在运动后24 h,CK活性再次升高到安静值的2.7倍。运动后72 h,CK活性再一次回到安静值水平。袁建琴等[49]研究发现,运动后骨骼肌损伤呈现延迟性发展的趋势,CK活性峰值出现在运动后即刻。于新凯等[50]研究发现,力竭性离心运动组大鼠CK峰值出现在运动后即刻,高出对照组6.67倍,随后逐渐下降,在7 d后虽然明显下降,但仍然高于正常值1.48倍。
本实验发现,运动后即刻血清CK活性比安静时升高6.93倍(P<0.01)。运动后12 h组血清CK活性比安静时升高3.71倍(P<0.01),运动后24 h组血清CK活性比安静时升高2.33倍(P<0.01),运动后48 h组血清CK活性比安静时升高2.06倍(P<0.01),运动后72 h组血清CK活性比安静时升高1.45倍(P>0.05),与上述研究结果基本一致。
目前,关于EIMI的机制学术界一直存在着不同的观点。主要有机械损伤说、细胞内钙稳态失调说和细胞骨架损伤说。我国学者认为,EIMI的原因,可能是骨骼肌细胞内Ca2+代谢紊乱,使细胞骨架蛋白发生了变化,如结蛋白等水解、解聚,从而使维护肌节结构稳定性的细胞骨架网络遭到破坏,导致骨骼肌超微结构的改变。Armstrong发现离心式运动可导致大鼠血清CK活性在运动后呈双相变化,即运动后酶活性迅速升高,6~12 h基本恢复正常,36 h后再次出现高峰,并且血清CK活性在第2次增加的程度更为显著。他认为造成这种现象的原因是由于EIMI引起部分肌纤维变性坏死、肌肉酶得以释放的结果。因此,较高的血清CK活性可能是骨骼肌纤维机械损伤的结果。尽管大量研究揭示了骨骼肌损伤与血清CK活性变化之间的可能关系,以及运动后血清CK活性变化的一些规律,但骨骼肌超微结构与血清CK活性对运动训练产生哪些适应性,尚需要进一步深入研究。EIMI既可能是一种主导因素引起的连锁反应,也可能是多种因素共同作用的结果。笔者认为,在非机械性损伤前提下,内环境失衡可能是导致EIMI的主要原因。不同机制下产生的EIMI需要更深一步的研究。
4 结论
1)大强度运动造成的大鼠骨骼肌超微结构损伤,与其他时刻相比,在运动后24 h程度最明显;之后逐渐得到恢复,72 h后恢复接近正常。
2)一次力竭性离心运动后即刻大鼠血清CK活性显著增高,随后逐渐恢复,72 h接近正常水平。运动损伤对CK的活性具有非常显著的影响。
3)骨骼肌超微结构变化与CK活性变化存在不同现象。CK的活性变化先于骨骼肌超微结构形态变化。CK的活性变化可以在一定程度上反应骨骼肌的损伤情况。
[1]Karoline Cheung.Delayed onset muscle soreness:treatment strategies and performance factors[J].Sports Med,2003,33(2):145-146.
[2]田野,马鹏鹏.连续运动后延迟性肌肉损伤的适应性研究[J].中国运动医学杂志,2003(22):138-142.
[3]李世成,焦海舟.大鼠离心运动后骨骼肌微细损伤机制的研究[J].湛江师范学院学报,2006,27(3):90-95.
[4]袁建琴,王瑞元.运动对结蛋白的影响[J].中国运动医学杂志,2006,25(2):210-212.
[5]罗吉伟,余斌,覃承诃,等.反复力竭运动后大鼠骨骼肌线粒体超微结构改变及维生素E的保护作用[J].第一军医大学学报,2003,24(12):1326-1328.
[6]于新凯,田野.下坡跑训练队大鼠血清CK、LDH的影响[J].上海体育学院学报,2000,8(3):29-32.
[7]Bedford T.G.,Tipton.,Wilson N.C.,et al.Maximum oxygen consumption of rats and its changes with various experimental procedures[J].Appl Physiol,1997(47):8750-7587.
[8]Friden J.Sublethal muscle fiber injuries after high-tension anaerobic exercise[J].Eur.J.Appl.Physiol,1988,57:360-368.
[9]Takekura H Nishizawa T Nishizawa T.Eccentric exercise-induced morphological changes in the membrane systems involved in excitation-contraction coupling in rat skeletal muscle[J].Journal of Physiology,2001,533:571-583.
[10]Armstrong RB.Initial events in exercise-induced muscular injury[J].Med Sci Sports Exerc,1990,22(4):429-435.
责任编辑:郭长寿
Different Exhausted Points After Eccentric Exercise Triceps Ultrastructure and Creatine Kinase Changes in Expression
SUN Liang,ZHAO Gang
(Jilin Normal University,Siping 136000,Jilin,China)
Through the establishment of sports injury model of rats after downhill running different exhausted points after the eccentric exercise and the ultrastructure of skeletal muscle injury mechanism were studied.The result is as follows:after acute eccentric exercise rat triceps ultrastructure changed,muscle cell membrane was fuzzy and dissolved,muscle fiber arrangement was in disorder,mitochondria exhibited swelling and vacuole degeneration,Z line was abnormal(included Zline flow,Z line vagueness,Z line twist and Zline disappearance),sarcoplasmic reticulum swelled and so on.Injury was most obvious 24 hours after exercise,and then restores gradually.It had basically restored 72 hours after exercise.The serum CK activity immediately after exercise(2035.42±426.49)U/L increased significantly compared with the control group(293.66±76.07)U/L(P<0.01)reached peak value,and then restored gradually,the serum CK activity 72 hours after exercise(425.51±143.34)U/L restored significantly compared with exercise group(P<0.01)24 hours after the eccentric exercise,72 hours after significantly restored.The conclusion is that an exhaustive centrifugal ultrastructural exercise rat skeletal muscle bodies changed,and the change was most obvious after 24 hours,followed by gradual resumption of exercise and the restore was significant after 72 hours.The first exhaustive exercise immediately after centrifugation serum CK activity was significantly increased and serum CK activity can reflect the changes in skeletal muscle ultrastructure after sports injury conditions.
exercise-induced skeletal muscle injury;skeletal muscle ultrastructure;creatine kinase
G804.7
A
1004-0560(2015)03-0074-05
2014-08-09;
2014-09-10
孙 良(1965—),男,副教授,学士,主要研究方向为体育教学与训练。
