福建三明栲类次生林主要树种更新生态位研究
2015-01-02郑小贤刘晓玥
周 洋,郑小贤,王 琦,刘晓玥
(北京林业大学 森林资源与环境管理国家林业局重点实验室,北京100083)
更新生态位最早由Grubb(1977)提出,主要指植物在更新过程中对生境条件、空间等生态因子的要求及适应,是植物更新和生态位研究的一个重要方面。通常指成年阶段具有相同生态位的树种之间的种子、幼苗和小树阶段的生态位分化[1-4]。更新生态位本身是一个动态的概念,但一直以来对更新生态位的测量都沿用生态位的测度公式,没有考虑到更新生态位的动态性[5-6]。因此,本研究把栲类次生林划分为幼苗层[7]:H<1m,幼树层:H>1m、DBH<5cm,小树层:5cm<DBH<10cm,大树层:DBH>10cm,结合态势理论[8-9],分别以幼苗层、幼树层、小树层、大树层的生态位作为更新生态位的态和势,第一更新层以幼苗层和幼树层生态位作为更新生态位的态和势,依次为第二和第三更新层,可以反映物种的更新趋势。
栲树(Castanopsis fargesii)是南亚热带常绿阔叶林中主要物种,其材质坚硬,色泽美观,具有较高的社会、经济、生态效益,以栲树为优势种组成的栲类次生林在福建三明地区有着广泛的分布,但由于长期以来对栲类次生林的不合理经营和人为干扰,造成群落物种多样性逐渐降低、目的树种丧失严重,形成大量的低质低效次生林,如何恢复和改造栲类次生林成为亟待解决的问题。有关栲类次生林的结构、地理分布、生态特征、功能和种群发展趋势等方面的研究已有较多的报道[10-15],但对栲类次生林树更新生态位的研究还较少。以福建三明将乐林场栲类次生林为研究对象,通过探讨其更新生态位,揭示在特定环境因子作用下的种间关系和演替中植物替代关系的机制,为福建三明栲类次生林生态恢复和经营提供科学依据。
1 研究地概况
研究区位于福建将乐县将乐林场,地处武夷山脉东南麓,平均海拔400~800m,为褶皱山地地貌。该地区位于中亚热带沿海海洋性季风气候,年平均气温18.7℃,极端最高气温41.1℃,极端最低气温-2.5℃,年平均降雨量1 204mm。土壤以红壤分布范围最广,土壤质地一般为沙壤土或轻壤土,块状结构,实度适中,腐殖质层厚度为4.5~15.5cm。林场占地面积7 113.6hm2,森林覆盖率为93.8%,森林类型以天然常绿阔叶次生林为主。常绿阔叶次生林乔木层主要树种有栲树(Castanopsis fargesii)、拟赤杨(Alniphyllum fortunei)、木荷(Schima superba)、甜 槠 (Castanopsis eyrei)、青 冈 (Cyclobalanopsis chungii)等。
2 材料与方法
2.1 取样方法
在2012年7月和2013年7月,采用典型取样法设置样地。设置栲类次生林固定样地18块,每个样地面积为20m×30m,每木检尺,记录乔木层胸径≥5.0cm的所有树种名称、胸径、树高、第一枝下高、冠幅;在样地4角及中心分别设置5个2m×2 m灌木样方和1m×1m草本样方,记录灌木层中植被名称、株数、高度,草本层(包括幼苗)中植物种类、数量、盖度、高度。样地基本情况见(表1)。
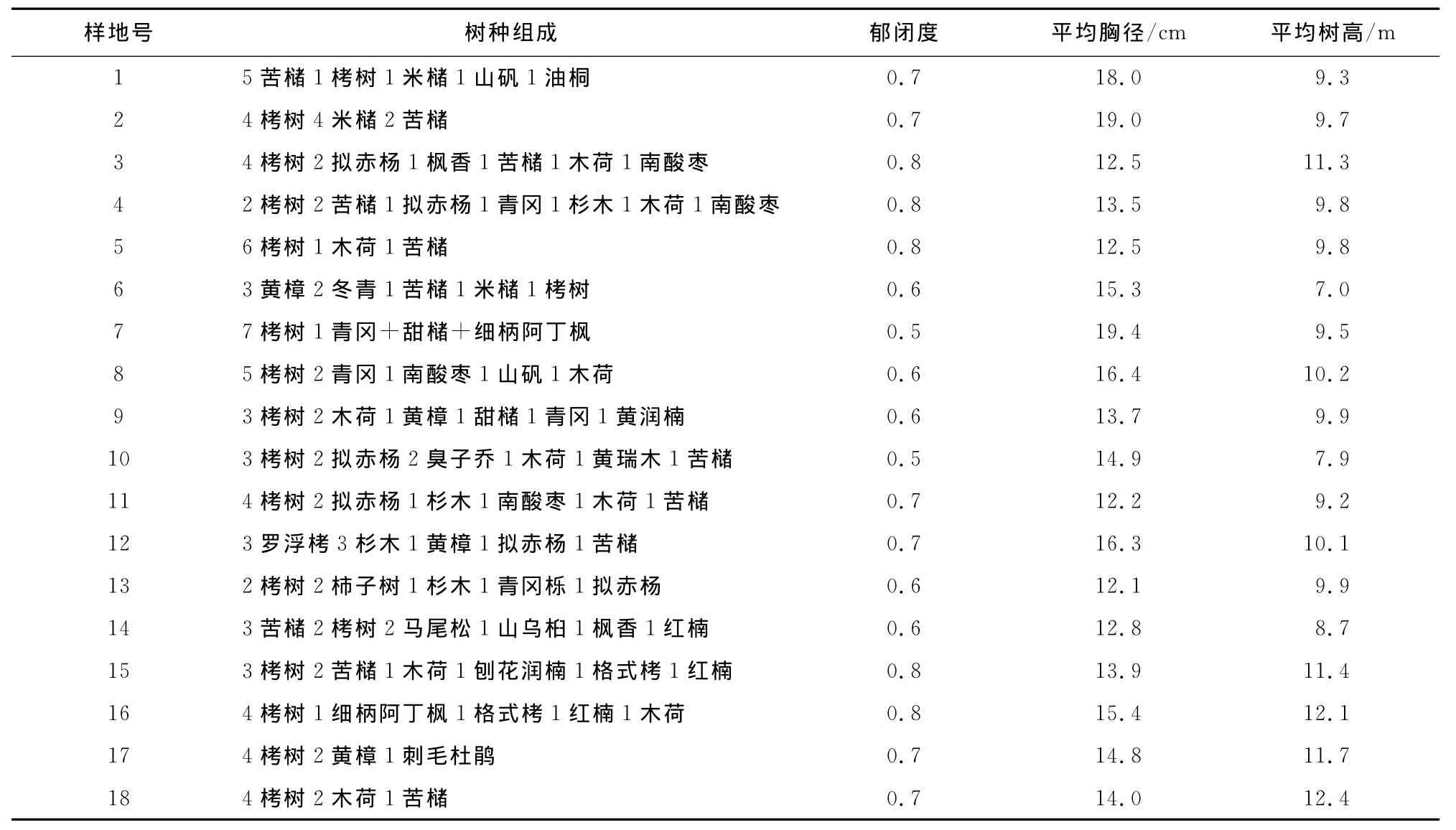
表1 样地基本情况Table 1 Basic situations of sample plot
2.2 改进的更新生态位宽度与生态位重叠测度
生态位宽度Shannon-Wiener指数和生态位重叠参照王琳[16]和 E.R.Pianka[5]的方法。改进的更新生态位宽度:以第一更新层即幼苗层到幼树层为例,改进的更新生态位宽度公式[8]:B'iB=(BiB+AiBis)/∑(BjB+AjBjs),式中:i,j=1,2,…n,B'iB为改进后的物种i的更新生态位宽度;BiB、BjB为物种i、j在幼树层的Shannon-Wiener生态位宽度值;Bis、Bjs为物种i、j幼苗层的Shannon-Wiener生态位宽度值;Ai、Aj为物种i、j在幼树层的重要值占所有幼树层重要值的百分比。改进的更新生态位重叠公式[17]:a'ij=(aB+aS)/2,式中:a'ij为改进后的更新生态位重叠值;aB和aS由Pianka指数计算得到,分别为幼树层和幼苗层的生态位重叠值。
2.3 树种重要值
树种重要值作为判断优势度的一个综合指标,能相对客观地反映树种在群落中的地位和作用,其计算公式为[18]:重要值(IV)=(相对密度+相对频度+相对显著度)/3。经过对栲类次生林群落调查数据的分析,选取前10位主要树种进行更新生态位分析(表2)。
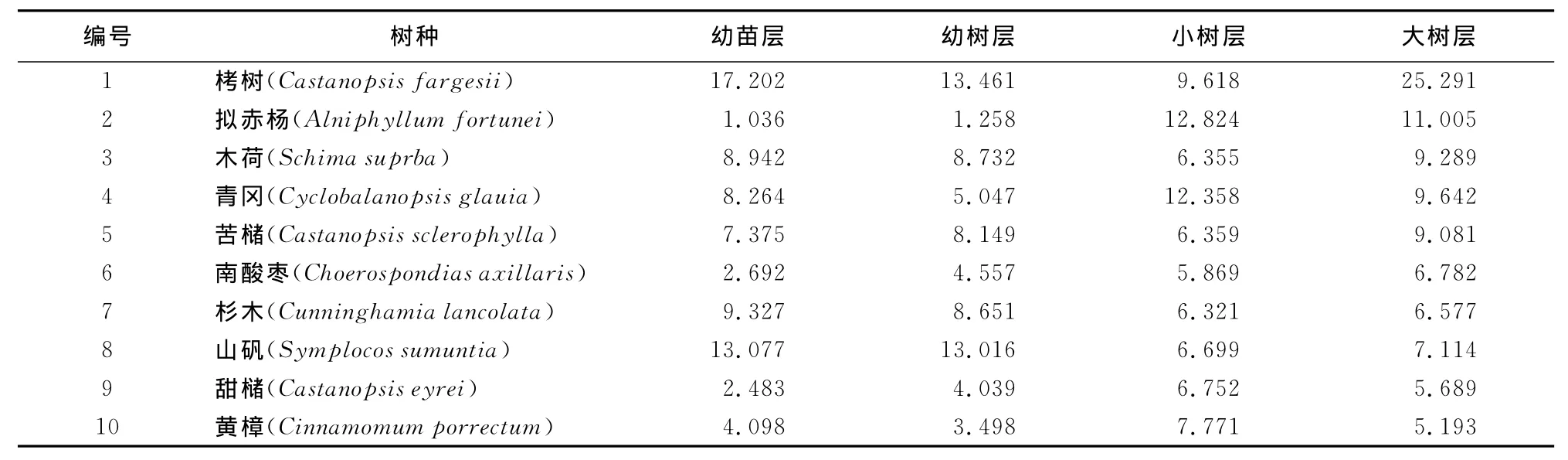
表2 各更新层主要树种的重要值Table 2 The important value of dominant species in each regeneration layers
3 结果与分析
3.1 栲类次生林主要树种重要值
栲类次生林中各层树种的重要值存在差异(表2),栲树处于优势地位,在幼苗层、幼树层、大树层中栲树的重要值都是最高。研究对象为1958年皆伐后自然恢复形成的栲类次生林,栲树为优势树种处在主林层。拟赤杨、木荷、青冈、苦槠在小树层和大树层的重要值都比较高,说明它们是栲树的主要伴生树种,但拟赤杨在幼苗层和幼树层的重要值很小,与其他树种在对空间资源利用的竞争中处于弱势。山矾在幼苗层和幼树层中重要值较大,处于优势地位,多见于林缘阴湿处,小树层和大树层中重要值较小,说明山矾幼苗较耐荫蔽,天然更新能力强,但在小树层和大树层中竞争不过栲树及其主要伴生树种。
3.2 各更新层主要树种的更新生态位
在第一更新层即幼苗层到幼树层过程中,栲树、木荷、青冈的更新生态位宽度位于前三(表3),都达到0.15以上,表明它们是在幼苗层到幼树层过程中天然更新能力强,能充分利用环境。相反,拟赤杨、南酸枣、杉木的更新生态位宽度较小,天然更新能力和竞争能力较弱,会随着演替的进行逐渐处于弱势。
第二更新层和第三更新层中,栲树、木荷、苦槠的更新生态位宽度均位于前三,甜槠、黄樟、杉木、山矾的更新生态位宽度较小,木荷、苦槠、青冈在林内分布较广、数量较多,在郁闭条件下,幼苗、幼树适应性能力较强,易与栲树形成混交林。虽木荷重要值不及栲树,但其更新生态位宽度值大,具有较大的生态适应范围,同时与栲树同在主林层,可以形成共优种。同栲树相比,苦槠和青冈的更新生态位宽度略小些,由于它们均为阳性常绿乔木,与栲树在同一林层冠幅高大,资源位利用有一定优势。其他树种如杉木、甜槠、黄樟的更新生态位宽度值较小,耐荫性差,在林内难以找到其幼苗、幼树,是群落中的衰退树种。南酸枣的更新生态位宽度呈上升趋势,主要分布在小树层,随着演替的进行其优势度有增大的趋势。
3.3 主要树种的更新生态位重叠
栲类次生林主要树种的更新生态位重叠值见表4。栲树与其他树种的更新生态位重叠值为0.15~0.63,并且栲树和青冈、拟赤杨和木荷、青冈和木荷,木荷和苦槠等的更新生态位重叠值均大于0.4,表明这些树种均能充分利用环境资源。更新生态位宽度较大的树种一般能产生较大的更新生态位重叠值,如栲树与木荷,青冈和木荷。栲树与更新生态位宽度较大的木荷、苦槠、青冈间有较大重叠值,表明栲树与它们具有相似的资源需求,在资源充分条件下可共享,在资源相对紧缺时存在竞争排斥的可能性。栲树对更新生态位宽度较窄的杉木产生较小的重叠值,反映出种间明显的异质性,难以产生共优状态,杉木在郁闭状态下难以自然更新,是逐步被替代的树种。更新生态位宽度较大的木荷对其他树种的重叠值都偏大,表明木荷易于与其他树种混交。
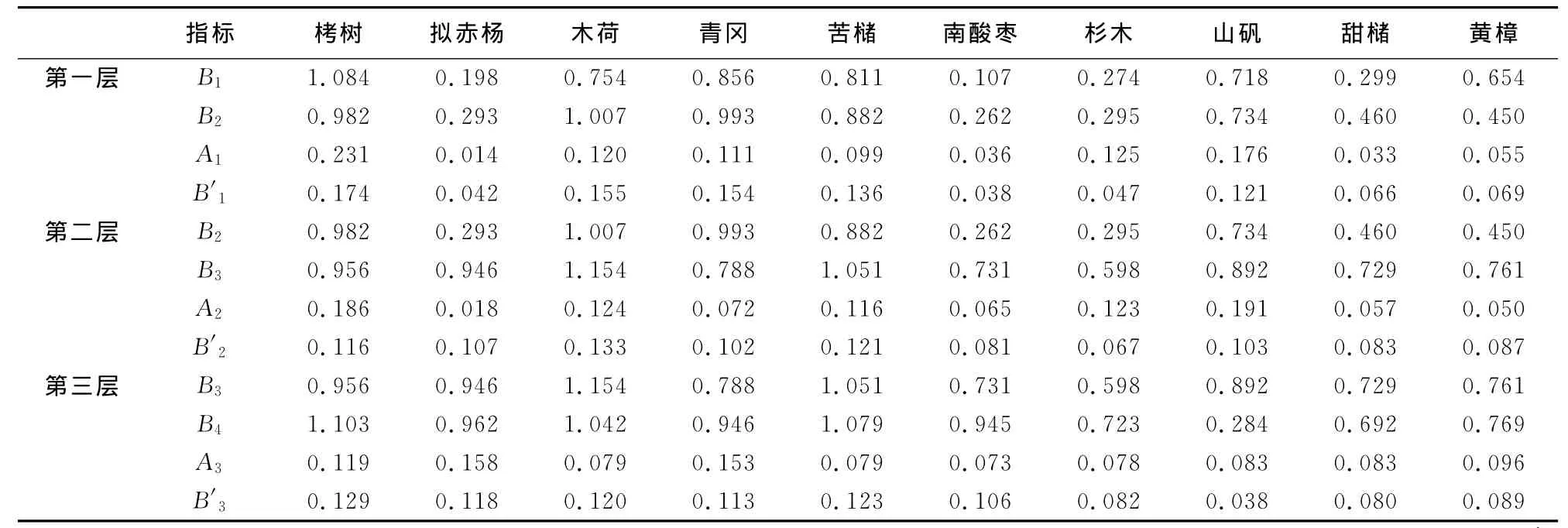
表3 主要树种更新生态位宽度值Table 3 Regeneration niche width of main trees

表4 主要树种的更新生态位重叠值Table 4 Regeneration niche overlap of main trees
4 结论与讨论
木荷、苦槠、青冈具有较大的更新生态位宽度,在林内分布较广、数量较多,利用资源较为充分,在郁闭条件下,幼苗、幼树适应性能力较强,易与栲树形成混交林。栲类次生林可能发展为以栲树、木荷、苦槠、青冈等树种为优势种的共优群落。拟赤杨虽然在小树层和大树层具有一定的优势度,但在幼苗层和幼树层重要值和更新生态位宽度都很低,表明与其他树种在对空间资源利用的竞争中处于弱势。
栲树、木荷、苦槠、青冈均具有较大更新生态位重叠值,它们具有相似的资源需求,对环境资源均能利用充分,在资源充分条件下可共享,在资源相对紧缺时存在竞争排斥的可能性。更新生态位宽度较大的树种一般能产生较大的更新生态位重叠值,如栲树与木荷,青冈和木荷。
甜槠、黄樟、杉木、山矾、杉木的更新生态位宽度较小,它们会随着演替的进行逐渐处于弱势,被其他树种替代。南酸枣的更新生态位宽度是上升趋势,主要分布在小树层,随着演替其优势度有增大的趋势。
物种更新生态位宽度主要体现树种利用资源能力的程度及生态适应性和分布幅度。在栲类次生林中,栲树重要值最大,更新生态位宽度也最大,表明栲树对资源利用率高,对环境的适应能力强,在维护群落结构和功能等方面,占据主要的生态位置,处于优势地位。木荷、苦槠、青冈具有较大的更新生态位宽度,幼苗、幼树适应能力较强,是栲树的主要伴生树种。甜槠、黄樟更新生态位宽度值较小,林内幼苗幼树较少,是群落中的衰退树种。杉木在郁闭状态下难以自然更新,是逐步被替代的树种。拟赤杨在幼苗层和幼树层的更新生态位宽度较低,在竞争中处于弱势,应采取人工抚育措施,促进拟赤杨幼苗幼树的天然更新。
更新生态位宽度较大的树种对资源利用能力较强,因而与其他树种间的生态位重叠较大,更新生态位宽度较小的物种对资源的利用能力较弱、因而与其他种群间的生态位重叠较小。具有较低生态位宽度的种群之间的生态位重叠并不一定较小。
[1] NAKASHIZUKA T.Species coexistence in temperate,mixed deciduous forests[J].Trends Ecol.Evol.,2001,16(4):205-210.
[2] GRUBB P J.The maintenance of species-richness in plant communities:The importance of the regeneration niche[J].Biol.Rev.,1977,52(1):107-145.
[3] 雷利平,王孝安,郭华,等.子午岭地区辽东栎和油松林建群种的更新生态位宽度分析[J].西北植物学报,2007,27(7):1446-1453.LEI L P,WANG X A,GUO H,et al.Dominant species of regeneration niche in Quercus Liaotungensis and Pinus tabulaeformis forest in Ziwuling Mountain[J].Acta Botanica Boreali-Occidentalia Sinica,2007,27(7):1446-1453.(in Chinese)
[4] 李永兵,王辉,付菁.子午岭辽东栎林林窗主要树种更新生态位[J].生态学杂志,2008,27(12):2062-2066.LI Y B,WANG H,FU J,Regeneration niche of main tree species in Quercus liaotungensis forest gaps in Ziwuling Mountain[J].Chinese Journal of Ecology,2008,27(12):2062-2066.(in Chinese)
[5] PIANKA E R.The structure of lizard communities[J].Ann.Rev.Ecol.Syst.,1973,4(1):53-74.
[6] 张光明,谢寿昌.生态位概念演变与展望[J].生态学杂志,1997,16(6):46-51.ZHANG G M,XIE S C.Development of niche concept and its perspectives[J].Chinese Journal of Ecology,1997,16(6):46-51.(in Chinese)
[7] 刘青松,齐代华,桂俊,等.四川山矾种群在不同群落中空间分布格局研究[J].西南农业大学学报,2006,28(3):418-421.LIU Q S,QI D H,GUI L,et al.Study on the distribution patterns of Symplocos setchuanensis population indifferent communities in the Jinyun Mountains[J].Journal of Southwest Agricultural University,2006,28(3):418-421.(in Chinese)
[8] 王莹莹,左金淼,刘家冈.以态势理论为基础的更新生态位测度研究 [J].林业科学,2005,41(4):20-24.WANG Y Y,ZUO J M,LIU J G.Study on regeneration niche metrics based on ecostate-ecorole theory [J].Scientia Silvae Sinicae,2005,41(4):20-24.(in Chinese)
[9] 朱春全.生态位态势理论与扩充假说[J].生态学报,1997,17(3):324-332.ZHU C Q.The niche ecostate-ecorole theory and expansion hypothesis[J].Acta Ecologica Sinica,1997,17(3):324-332.(in Chinese)
[10] 范海兰,洪伟,吴承祯,等.福建大田栲树次生林优势种群结构与动态[J].植物资源与环境学报,2004,13(3):44-49.FAN H L,HONG W,WU C Z,et al.Population structure and dynamics of dominant species in Castanopsis fargesii secondary forest in Datian County,Fujian Province[J].Journal of Plant Resources and Environment.2004,13(3):44-49.(in Chinese)
[11] 刘益曦,刁军.浙江天然常绿阔叶林空间结构特征分析[J].西北林学院学报,2014,29(4):155-159.LIU Y X,DIAO J.Spatial structural characteristics of natural evergreen broad-leaved forest of Zhejiang[J].Journal of Northwest Forestry University,2014,29(4):155-159.(in Chinese)
[12] 刘金福.格氏栲种群结构与动态规律研究[D].北京:北京林业大学,2004.
[13] 邓贤兰.江西井冈山栲属群落生态学特征研究[D].重庆:西南师范大学,2002.
[14] 吴大荣,苏志尧,李秉滔,等.福建三明莘口青钩栲种群结构和空间分布格局动态初步研究[J].林业科学,2000,36(3):27-32.WU D R,SU Z Y,LI B T,et al.Preliminary study on the structure and spatial pattern dynamics of Castanopsis kawakamii population in Xinkou Nature Reserve,Sanming of Fujian[J].Scientia Silvae Sinicae,2000,36(3):27-32.(in Chinese)
[15] 杜燕,郑小贤,高祥,等.森林物种多样性保育价值评价方法改进-以将乐林场栲树次生林为例[J].西北林学院学报,2013,28(4):176-179.DU Y ,ZHENG X X,GAO X,et al.Improvement of evaluation method of forest species diversity conservation value-a case study of Jiangle County[J].Journal of Northwest Forestry University,2013,28(4):176-179.(in Chinese)
[16] 王琳,张金屯.濒危植物矮牡丹的生态位研究[J].生态学杂志,2001,20(4):65-69.WANG L,ZHANG J T.The niche characteristics of endangered plant paeonia Suffruticosa var.Spontanea [J].Chinese Journal of Ecology,2001,20(4):65-69.(in Chinese)
[17] 陈志明,伍国仪,缪绅裕,等.大东山长柄双花木群落主要种更新 生 态 位 研 究 [J].安 徽 农 业 科 学,2013,41(28):11414-11416.CHEN Z M,WU G Y,MIU S Y,et al.Studies on the regeneration niche of main species in Disanthus cercidifolius var.longipes community in Dadongshan Mountain[J].Journal of Anhui Agricultural Sciences,2013,41(28):11414-11416.(in Chinese)
[18] 宋永昌.植被生态学[M].上海:华东师范大学出版社,2001:143-156.
