伏牛山自然保护区森林生态系统乔木层优势种种群动态
2015-01-02范玉龙丁圣彦卢训令
范玉龙,胡 楠,丁圣彦,卢训令
(1.河南大学 环境与规划学院,河南 开封475001;2.南阳理工学院,河南 南阳473004)
伏牛山国家级自然保护区是中国东部森林样带中亚热带和暖温带的结合点,低海拔山区是典型的落叶阔叶林幼林区,随着海拔的升高,人类活动影响减小,森林的原始状态逐渐显现,特别在高海拔区域,保存着完整的原始成熟林。区内乔木层占绝对优势,主导着森林生态系统植物群落的外貌、结构和功能。研究伏牛山森林生态系统乔木主要优势种组成及种群动态,可以了解次生落叶阔叶林在发育过程中物种的变化,也是对自然保护区天然林禁伐成果的检验。
地理位置特殊的伏牛山自然保护区一直是森林研究的热点区域。尚富德[1]等对伏牛山南北自然过渡地带植物多样性的特征及其成因进行了分析;张建伟、曾颖[2]分析了伏牛山区植物的多样性及其保护对策;胡楠、范玉龙[3-8]等对伏牛山自然保护区森林生态系统进行了详细的野外调查,在此基础上进行了植物功能群划分。树种在与环境因子相互作用过程中形成了较好的适应关系,树种分布格局对环境因子的空间结构形成了空间依赖性[9],不同演替阶段的树种种类及其重要性都发生着变化[10]。在伏牛山自然保护区,从低山丘陵的灌丛到山顶保护完好的成熟林,栎类群落变化明显。尽管栎林研究备受关注[11-16],但以径级结构为主分析其种群动态的还比较少,个体高度及胸径的变化可以更为简明地反映出种群动态变化。
1 研究地概况
伏牛山自然保护区位于河南省西南部,地理坐标为110°30′-113°05′E,32°45′-34°00′N,伏牛山地处暖温带向北亚热带过渡地带,植被属暖温带落叶阔叶林向亚热带常绿阔叶林的过渡型。伏牛山区的土壤复杂多样,可划分为3个土类(山地棕壤土类、山地黄棕壤土类和山地褐土土类)[1]。伏牛山还是长江、黄河、淮河3大水系一些支流的发源地,为重要的水源涵养林区。伏牛山北坡的洛阳白云山和南坡的南阳宝天曼自然环境随海拔变化差异显著,优势种变化明显,且乔木层在整个山体上呈现明显的地带性分布(表1)。

表1 研究区自然地理概况Table 1 Climate and geographical conditions in the experimental sites
2 材料与方法
2.1 样方设置
从南坡的宝天曼到北坡的白云山,在海拔600 m至2 000m的山体上,选择具有代表性的地段,用群落生态学的方法调查伏牛山自然保护区森林生态系统。每隔海拔100m确定至少1个20m×20m的样地,共设置样地66个[3]。
2.2 调查方法
用群落生态学的样方法调查研究区主要森林生态系统类型特征。调查指标包括:乔木层的郁闭度、种类、个体数量、树高、枝下高、冠高、冠幅、基径、胸径等[3]。在群落调查的基础上,通过计算重要值等方法,分析研究区各层优势种随海拔的分布状况。
2.3 径级结构划分方法
由于研究对象为乔木,年龄调查受到方法上的限制,野外调查年轮木芯难于钻取,考虑到本研究实际情况,结合高贤明[17]等的方法,采用径级、高度级来代表乔木树种的年龄结构。划分的标准为:更新层分幼苗为SC 1(幼苗,H≤50cm)、SC 2(幼树,50 cm<H≤100cm)、SC 3(大幼树,H>100cm,DBH<4cm)3个级别;然后根据树木的胸径,从4cm开始,胸径每增加4cm为1个径级,即SC 4(4cm≤DBH<8cm)、SC 5(8cm≤DBH<12cm)、SC 6(12cm≤DBH<16cm)、SC 7(16cm≤DBH<20 cm)、SC 8(20cm≤DBH<24cm)、SC 9(24cm≤DBH<28cm)、SC 10(DBH≥28cm)。
高度级划分标准:Ⅰ级(0m≤H<3m)、Ⅱ级(3m≤H<6m)、Ⅲ级(6m≤H<9m)、Ⅳ级(9m≤H<12m)、Ⅴ级(12m≤H<15m)、Ⅵ级(15m≤H<18m)、Ⅶ级(18m≤H<21m)。
3 结果与分析
通过对植被调查结果的分析发现,乔木优势种分布在南、北坡随海拔梯度呈现规律性变化(表2、表3),以壳斗科的植物为主的栓皮栎、短柄枹、锐齿栎及松科华山松等构成了保护区森林生态系统的建群种,这些建群种控制了伏牛山森林生态系统的外貌、结构和功能。
在北坡白云山,海拔1 000m以下以栓皮栎为绝对优势种,最大的伴生种为槲栎;海拔1 000~1 800m的优势种为锐齿栎,伴生种随海拔升高依次是短柄枹和华山松等;海拔1 800m以上以华山松为优势种。在南坡宝天曼,海拔1 200m以下栓皮栎是优势种;海拔1 200m上下短柄枹分布比较多;海拔1 200m以上以锐齿栎为优势种。从整个山体上来看,海拔1 000m以下以栓皮栎为主要的建群种;海拔1 200m以上以锐齿栎为主要的建群种;海拔1 400m以上的局部地段和海拔800~1 500m之间部分地段分别有以华山松和油松为主的群落;海拔1 400m以上,华山松开始出现,并与锐齿栎争夺空间,某些地段华山松成为优势种,但大部分地带还是以锐齿栎为优势种;短柄枹的分布随海拔呈抛物线样,从海拔600m到1 600m均有分布,海拔600~1 400m为其主要分布区域,海拔1 200m左右分布数量较多,在有些地段成为优势种[3]。
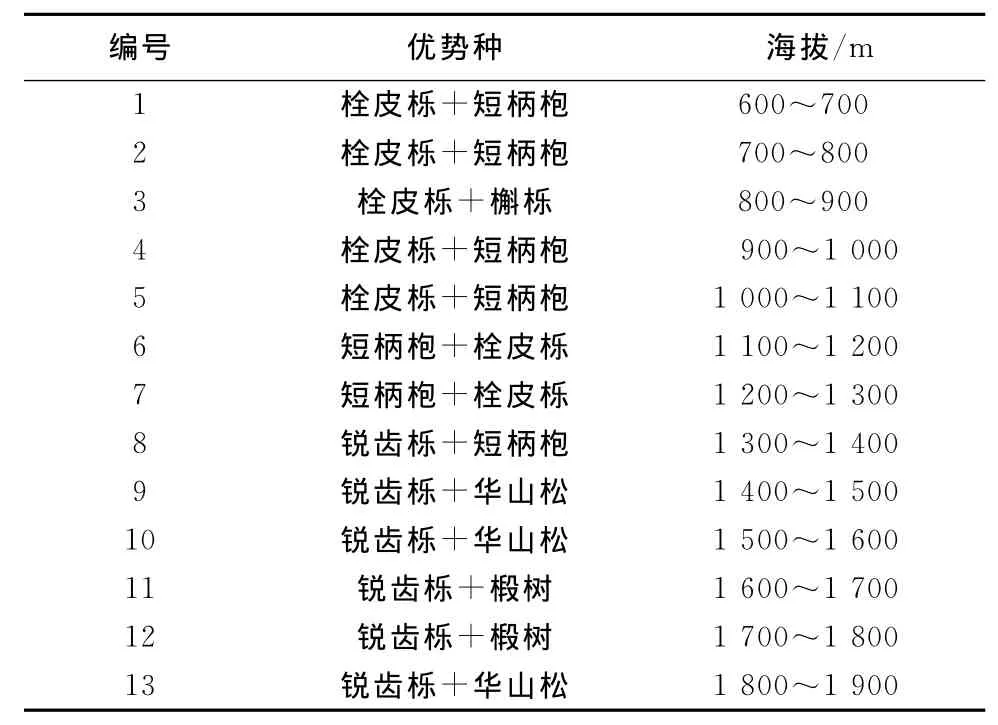
表2 南坡乔木优势种组合随海拔分布情况Table 2 The distribution of tree dominant species combinations along the altitudinal gradient on the south slope
3.1 南坡优势种的径级、高度级特征
南坡各优势种多呈现为稳定型,中龄个体较多,幼、老龄个体较少,天然更新良好,种群维持着一定的后续资源。在高海拔地段,优势种种群表现为中老龄个体较多,种群密度大,因该地段人为干扰较少,从而保存较多老龄个体,可以理解为正向演替的顶级阶段;中海拔地段,锐齿栎种群表现为缺少中、幼龄个体,老龄个体较多,种群缺乏后备资源的势态,该地段属锐齿栎种群的生存下限,种群密度小且波动性大(图1)。
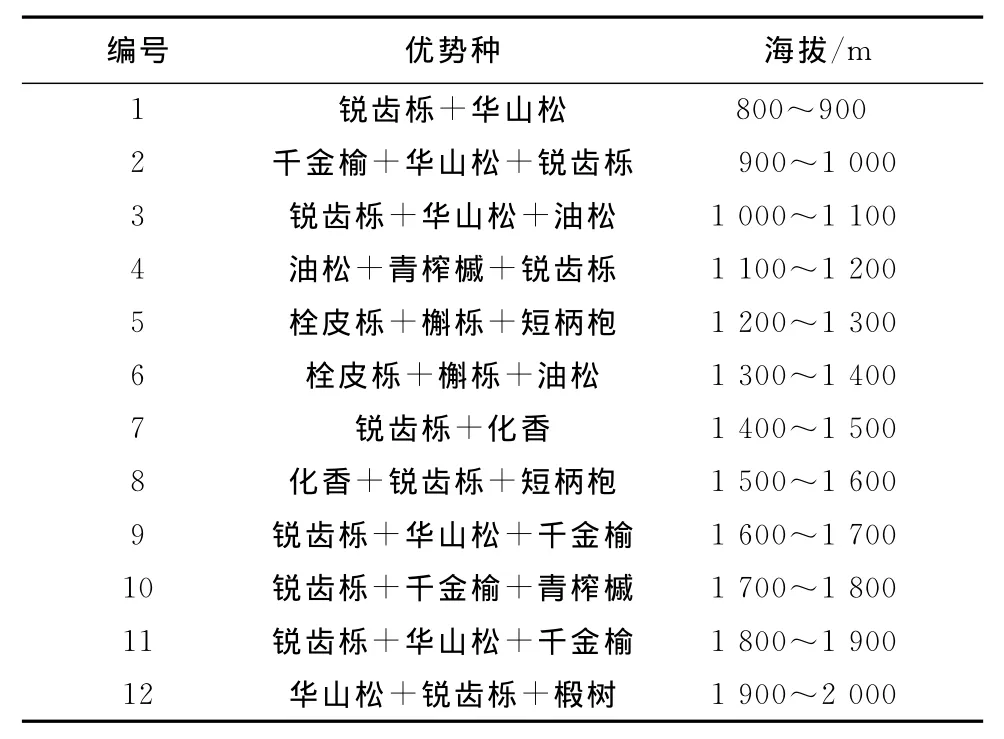
表3 北坡优势种组合随海拔分布Table 3 The distribution of dominant species combinations along the altitudinal gradient on the north slope

图1 南坡优势种径级结构随海拔变化Fig.1 Size class of the dominant species along altitude change at the south slope
在伏牛山南坡低海拔区域(600~1 000m),大量分布着栓皮栎和短柄枹的幼树,以高度级为Ⅲ级(树高为6~9m)的树木为主;在中海拔地段(1 000~1 200m),栓皮栎仍以高度级为Ⅲ级的树木较多,但是数量已经明显下降,而高度级为Ⅲ级的短柄枹数量显著增加,说明此处短柄枹正取代栓皮栎成为优势种,在个别地带,存在少量的锐齿栎大树(高度级为Ⅵ级);在高海拔地段(1 200~1 700m),短柄枹的大树比例明显增加,但是数量减小,而锐齿栎大树(高度级为Ⅵ级)数量已经占优势;在山顶(1 700~2 000m),大量分布着锐齿栎和华山松的混交林,它们的龄级分布比较广泛,说明它们在这个地段的更新及生长良好(图2)。
值得注意的是,在中海拔地段(1 000~1 200 m),有锐齿栎大树出现,但是中幼龄的锐齿栎却没有出现,只有少量幼苗,说明锐齿栎在这个地段的更新能力很弱,会逐渐消失,但是它们为什么在这个地段保留着少量大树,是否因为气候变化使锐齿栎整体上移所造成的,这还需要进一步的研究。

图2 南坡优势种数量的高度级分布Fig.2 Individual number vs height distributions of dominant species at the south slope
3.2 北坡优势种的径级结构特征
北坡优势种种群的径级结构特征与南坡基本相似,但同时也存在一些差异性,在低海拔地区,栓皮栎、短柄枹种群的幼龄个体比例明显低于南坡。在中海拔地区,北坡锐齿栎种群的种群密度远大于南坡(图3、图4)。北坡除气候因素外,人为因素也较南坡有较大不同,加上北坡很多地方坡度较南坡陡,使植被被破坏的可能性降低,这也是北坡与南坡植被差异的原因之一。
在伏牛山北坡低海拔区域(600~1 000m),栓皮栎和短柄枹的树高明显较南坡高,并且在这个海拔已经出现锐齿栎的大树;在中海拔地段(1 000~1 200m),短柄枹、锐齿栎占明显优势,特别是锐齿栎的幼树和大树均有大量分布,更新良好;在高海拔地段(1 200~1 700m),短柄枹几乎没有出现,锐齿栎和华山松的混交林已经占优势;在山顶(1 700~2 000m),锐齿栎的更新及生长良好(图4),这和径级结构反映的情况是一致的。
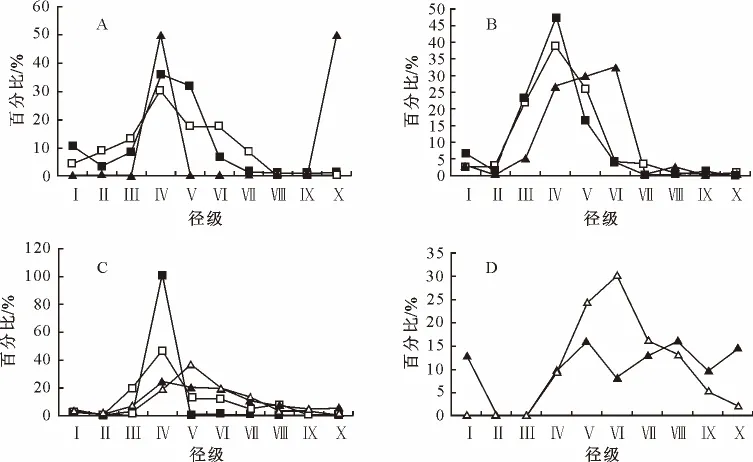
图3 北坡海拔变化优势种的径级结构Fig.3 Size class of the dominant species along altitude change at the north slope
群落自身相互作用、人类采伐经营活动以及一些未涉及的环境因子、干扰事件或随机因子也影响着植物种群的空间格局[18]。伏牛山自然保护区位于农业发达、人口众多的河南省,森林生态系统受人为破坏较为严重,特别是在低海拔区域,村庄分布较多,较大的树木基本不存在,中高海拔区域虽然无村落分布,但也受到了不同程度的影响,在高海拔区域,人为影响较少,保存着完整的原始天然次生林。随着海拔的升高,森林生态系统处于不同的发育期,群落结构发生着明显的变化,从某种程度上来说,这种变化代表了伏牛山自然保护区森林生态系统的不同演替阶段。虽然建群种也随海拔发生了变化,但是整体上还是从幼林到成熟林的渐变。天然林保护工程实施以来,自然保护区得以得到全面的休整,人工种植和自然恢复连续进行,使得种群朝着自然状态发展。优势种径级结构和树木高度的变化趋势说明:从低海拔到高海拔特别是在中低海拔,以前破坏严重的森林生态系统得到了恢复。除乔木层外,林下的灌木和草本层也得以恢复,整个森林生态系统的结构和功能朝着顶极阶段发展。在不同的地段,优势树种也发生着变化,随着天然林保护时间的增加,一些先锋种和保留种的更替也不断地进行,整个森林生态系统变得更加稳定和多样性。但是人工林及半自然状态下的森林生态系统中缺乏原始林内的树种,可能是由于森林植物的扩散限制[19],更深层的原因还需要进一步研究。
4 结论与讨论
伏牛山自然保护区森林生态系统乔木层优势种明显,主要的建群种随海拔梯度依次是栓皮栎、短柄枹、锐齿栎、华山松,它们控制着整个山体的外貌与结构。在个别地带的优势种会发生局部变化,如椴树、千金榆和三桠乌药等会在个别地段成为优势种。以樟科为代表的植物在沟谷地段成为优势植物,说明伏牛山自然保护区植被属含常绿成分的落叶阔叶林。
优势种种群动态在一定的海拔地段多呈现为稳定型,而在其他地段它们的更新能力较弱,说明不同优势种有着各自的适应范围。整体上,伏牛山自然保护区森林生态系统中龄个体较多,幼、老龄个体较少,天然更新良好,种群维持着一定的后续资源;在高海拔地段,优势种种群表现为中老龄个体较多,幼龄个体较少,种群处于正向演替的顶级阶段;在低海拔地段,优势种种群表现为中、幼龄个体较多,老龄个体较少,种群处于正向演替初期阶段。天然林禁伐后,伏牛山自然保护区森林生态系统得到了充分的保护,森林的自我修复使得森林生态系统的结构更加完整。在以前人为破坏较大的区域,人工造林和自然恢复使这部分森林迅速的发育。并且在研究中发现,在森林快速发育阶段,增长最快的是优势种,它们迅速占据空间,群落郁闭度高,林下灌木和草本植物较少,随着优势种的进一步发育,郁闭度有所降低,林下灌木和草本植物才渐渐丰富起来,整个
群落的植物多样性开始增加。
由于人为活动的影响,伏牛山自然保护区的海拔梯度变化可以为森林生态系统的不同演替阶段提供参考,为恢复破损的森林生态系统提供依据。
在不同海拔,乔木优势种的径级和高度级基本是吻合的。优势种在海拔梯度上的动态变化反映出小气候变化对树种分布的影响,南北坡优势种随海拔变化的差异也反映出纬度变化对植被分布的影响。

图4 北坡优势种数量的高度级分布Fig.4 Individual number vs height distributions of dominant species at the north slope
木本植物的径级系列可视为年龄的一种等级系列,二者具有同等的连续性含义[20-21]。因此,个体的高度及胸径变化能更好地反映出种群动态[17]。
研究种群的径级结构可以反映种群的稳定性[22]、种群的数量动态[23]、种群与环境的相互关系及其在群落中的地位和作用[21],能较好地反映出伏牛山自然保护区落叶阔叶林乔木层优势种组成及其种群动态。从种群动态的微小变化中,寻找自然规律[24-25],可以预测气候变化对优势种分布的影响,这也为我们在全球气候变化的背景下寻找植被响应规律提供直接的方法。
[1] 尚富德,王磐基,冯广平,等.伏牛山南北自然过渡地带植物多样性的特征及其成因分析[J].河南大学学报:自然科学版,1998,28(1):54-60.SHANG F D,WANG P J,FENG G P,et al.Study on the characteristics and causes of formation of plant diversity in the Funiushan Transition Region[J].Journal of Henan University:Natural Science,1998,28(1):54-60.(in Chinese)
[2] 张建伟,曾颖.伏牛山区植物的多样性及其保护[J].河南大学学报:自然科学版,2000,30(1):76-81.
[3] 胡楠,范玉龙,丁圣彦,等.伏牛山自然保护区森林生态系统乔木植物功能型分类[J].植物生态学报,2008,32(5):1104-1115.
[4] 范玉龙,胡楠,丁圣彦,等.伏牛山自然保护区森林生态系统草本植物功能群的分类[J].生态学报,2008,28(7):3092-3101.FAN Y L,HU N,DING S Y,et al.A study on the classification of plant functional types based on the dominant herbaceous species in forest ecosystem at Funiu Mountain National Natural Reserve[J].Acta Ecologica Sinica,2008,28(7):3092-3101.(in Chinese)
[5] 胡楠,范玉龙,丁圣彦,等.伏牛山森林生态系统灌木植物功能型分类[J].生态学报,2009,29(8):4017-4025.HU N,FAN Y L,DING S Y,et al.A study on the shrub classification of plant functional groups in forest ecosystem at Funiu Mountain[J].Acta Ecologica Sinica,2009,29(8):4017-4025.(in Chinese)
[6] 胡楠,范玉龙,丁圣彦,等.伏牛山自然保护区森林土壤与植物功能群关系研究[J].河南大学学报:自然科学版,2009,39(1):72-76.
[7] 胡楠,范玉龙,丁圣彦,等.伏牛山自然保护区北坡主要乔木优势种光合作用特性研究[J].河南大学学报:自然科学版,2009,39(4):392-396.HU N,FAN Y L,DING S Y,et al.Study of tree layer dominant species photosynthetic characteristics at north slope of Funiu Mountain Nature Reserve[J].Journal of Henan University:Natural Science,2009,39(4):392-396.(in Chinese)
[8] 胡楠,范玉龙,丁圣彦,等.伏牛山自然保护区不同演替期乔木优势种空间分布格局比较[J].河南大学学报:自然科学版,2010,40(5):493-497 HU N,FAN Y L,DING S Y,et al.Spatial distribution patterns of dominant species at different successional phase at Funiu Mountain National Nature reserve[J].Journal of Henan University:Natural Science,2010,40(5):493-497.(in Chinese)
[9] DRAY S,LEGENDRE P,PERES-NETO PR.Spatial modelling:a comprehensive framework for principal coordinate analysis of neighbour matrices(PCNM)[J].Ecological Modelling,2006,196:483-493.
[10] 张春雨,赵秀海,赵亚洲,等.长白山温带森林不同演替阶段群落结构特征[J].植物生态学报,2009,33(6):1090-1100.ZHANG C Y,ZHAO X H,ZHAO Y Z.Community structure in different successional stages in north temperate forests of Changbai Mountains,China[J].Acta Phytoecologica Sinica,2009,33(6):1090-1100.(in Chinese)
[11] SORK V,BRAMBLE SEXTON.Ecology of mastfruiting in three species of Missouri oaks,Quercus alba,Quercusrubra,and Quercus velutina (Fagaceae)[J].Ecology,1993,74:528-541.
[12] KOENIG W,MUMME W,CARMEN.A corn production by oaks in central coastal California:variation within and among years[J].Ecology,1994,75:99-109.
[13] FOX J.Adaptation of gray squirrel behavior to autumn germination by white oak acorns[J].Evolution,1982,36:800-809.
[14] ANDERSSON C.The effect of weevil and fungal attacks on the germination of Quercus robur acorns[J].Forest Eco.logy and Management,1992,50:247-251.
[15] NEGI A,NEGI G,SINGH S.Establishment and growth of Quercus floribundaseedlings after a mast year[J].Journal of Vegetation Science,1996,7:559-564.
[16] MATSUDA K.Seedling and growth of oak(Quercus serreta Thunb.)seedlings in an abandoned coppice forest[J].Ecological Research,1989,4:309-321.
[17] 高贤明,王巍,杜晓军,等.北京山区辽东栎林的径级结构、种群起源及生态学意义[J].植物生态学报,2001,25(6):673-678.GAO X M,WANG W,DU X J,et al.Size structure,ecological significance and population origin of Quercus wutaishanica forest in Beijing mountainous area [J].Acta Phytoecologica Sinica,2001,25(6):673-678.(in Chinese)
[18] BORCARD D,LEGENDRE P,DRAPEAU P.Partialling out the spatial component of ecological variation[J].Ecology,1992,73:1045-1055.
[19] PERTERKEN G.A met hod for assessing woodland flora for conservation using indicator species[J].Biological Conservation,1974(6):239-245.
[20] 徐学红,于明坚,胡正华.浙江古田山自然保护区甜槠种群结构与动态[J].生态学报,2005,25(3):645-653.XU X H,YU M J,HU Z H,et al.The structure and dynamics of Castanopsis eyrei population in Gutian Mountain Natural Reserve in Zhejiang,East China[J].Acta Ecologica Sinica,2005,25(3):645-653.(in Chinese)
[21] 张光富.浙江天童灌丛群落中优势种群的年龄结构和分布格局[J].武汉植物学研究,2001,19(3):233-240.ZHANG G F.Studies on the age structure and spatial pattern of dominant populations from Tiantong shrub community in Zhejiang Province[J].Journal of Wuhan Botanical Research,2001,19(3):233-240.(in Chinese)
[22] MANUEL C.Ecology:Concepts and application[M].Beijing:Science Press,2000:162-226.
[23] HURLBERT S.The measurement of niche overlap and some relatives[J].Ecology,1978,59(1):67-77.
[24] 曹小玉,李际平.福寿林场杉木人工林林下植物物种多样性研究[J].西北林学院学报,2014,29(3):57-61.CAO X Y,LI J P.Species diversity of undergrowth vegetation of Chinese fir plantations in Fushou Forest Farm [J].Journal of Northwest Forestry University,2014,29(3):57-61.(in Chinese)
[25] 张建华,丁国栋,姜鹏,等.沿坝生态交错带不同落叶松群落生长动态研究[J].西北林学院学报,2014,29(2):42-48.ZHANG J H,DING G D,JIANG P,et al.Growth dynamic of larix principis-rupprechtii Mayr.community along the dam ecotone[J].Journal of Northwest Forestry University,2014,29(2):42-48.(in Chinese)
