祁连山北坡中部祁连圆柏种群空间分布格局
2015-01-02占玉芳
占玉芳
(张掖市林业科学研究院,甘肃 张掖734000)
自从1952年Greig-Smith首次使用植物种群格局规模分析方法研究植物种群的格局以来,在这方面取得了重大进展[1-2],目前格局分析已成为生态学的研究热点之一。从20世纪80年代以来,我国在植物种群分布格局方面研究较多[2]。空间分布格局是植物种群研究的重要内容,是种群的重要结构特征之一[3]。植物种群空间分布格局,是关于种群个体间在水平空间上的配置状况或分布状况,也反映了种群个体与环境的相互关系及其在群落中的作用和地位[4-5]。种群空间分布格局一般分为随机分布、聚集分布和均匀分布3种类型[6]。不同的格局类型反映种群利用环境资源的状况,并可以揭示种群生物学内涵,是种群在群落中地位与综合生存能力的外在表现[7],因而种群空间分布格局通常反映了种群个体行为、生存和生长对一定环境因子的响应[8-9]。研究濒危物种种群空间分布格局,既可进一步了解群落空间结构特征和对种群的水平结构进行定量描述,也可阐明濒危物种种群及群落的动态变化,为制定其保护措施提供科学理论依据[10-11]。
祁连圆柏(Sabina przewalskii),是中国特有的常绿针叶树种,一般高达12m,常绿乔木,原产于四川北部、甘肃河西走廊及南部、青海,以它为建群种的天然林广泛分布于中国西北部海拔2 600~4 000 m的亚高山及高山地区[12]。祁连圆柏具有耐寒、耐旱和抗盐碱等特性,对于维系祁连山森林生态系统生物多样性、涵养水源、保持水土具有非常重要的生态作用。该群落主要占据阳坡或半阳坡形成疏林,并和位于阴坡的青海云杉一起形成具有两大特色的群落类型。此外,通过所得年轮数据发现,祁连圆柏树龄高达1 800a余,是中国目前已知可用来在干旱、半干旱区建立上千年甚至2 000a以上年表的唯一天然树种,对了解过去千年气候变化具有重要作用[13]。很多研究表明,不同生境下祁连圆柏对气候因子的响应存在一定的差异,已对其树轮生态学[14]、生理生态学[15]等方面进行了研究,但对祁连圆柏天然林空间分布格局很少报道。本研究对集中分布区祁连圆柏种群进行了野外调查,采用方差均值比方法判定分布格局类型,分析了不同尺度及不同海拔高度祁连圆柏种群空间分布格局,旨在揭示珍稀树种祁连圆柏种群分布特征及空间结构,进一步了解种群数量动态和发展趋势,为探索祁连圆柏种群濒危机制、阐述种群的空间动态及合理保护和经营利用祁连圆柏资源提供依据,并为祁连圆柏群落生态学的深入研究,特别是祁连山阳坡的植被恢复提供一定的理论基础。
1 研究区概况
位于甘肃省张掖市肃南县寺大隆林区的天涝池流域,地理坐标99°31′-100°15′E,38°14′-38°44′N,海拔高度在2 500~3 600m之间,年平均气温0.7℃,年降水量440.58mm,年蒸发量1 061.8 mm,年均相对湿度60%,属温带高寒半干旱气候。土壤主要是山地森林灰褐土。土壤和植被随山地地形和气候的差异而形成明显的垂直分布带。青海云杉(Picea crassifolia)为唯一建群种,面积占水源林面积的24.74%,占祁连山乔木林总面积的75.72%,呈斑块状分布于阴坡、半阴坡;祁连圆柏(Sabina przewalskii),呈小块状分布于阳坡、半阳坡;因生长环境条件的限制,乔木层比较单一,很少有其他伴生树种。灌木层常见的植物有金露梅(Dasiphora fruticosa)、箭 叶 锦 鸡 儿 (Caragana jubata)、刚毛忍冬(Lonicera hiapida)、高山绣线菊(Spiraea alpina)等,植被覆盖度约为40%。植物以禾本科植物为主,数量较多的有垂穗披碱草(Elymus nutans)、短颖鹅观草(Roegneria breviglumis)等,其次为大披针苔草(Carex lanceolata)、红棕苔草 (C.digyna)、北方嵩草 (Kobresia bellardii)、矮嵩草(K.humilis)、以及绢茸火绒草(Leontopodium smithianuin)、高山唐松草(Thalictrum alpinum )、宽苞棘豆(Oxytropis latibracteata)等。
2 材料与方法
2.1 野外调查及数据记录
于2012年7月,在不同海拔梯度天然祁连圆柏林群落中,采用样地法结合样方法对祁连圆柏自然种群进行调查,设置2 800、3 000、3 200m3个不同海拔梯度,每个海拔高度各设置3个20m×20m的样地,共建立调查样地9个。在每个样地中利用网格样方法将其分割成16个5m×5m的小样方,在每个小样方内对所有所有祁连圆柏进行每木调查,乔木和灌木记录特种名称、高度、胸径或地径、冠幅等。以每个样地的2个垂直边为坐标轴,记录每株植物的坐标(x,y),对祁连圆柏进行定位。同时,在每个小样方中心设置1m×1m草本样方1个,调查乔木及灌木幼苗记录特种名称、高度、地径,草本记录特种名称、株高、株数、盖度等。同时记录每个样地的地理位置、乔木层盖度、灌木层盖度、草本层盖度、海拔、坡度、坡向、土层厚度、凋落物厚度及人类活动的干扰情况等环境因子。
2.2 分布格局类型的测定
以样地内祁连圆柏个体定位数据为依据,应用相邻格子法,分5m×5m、5m×10m、10m×10m的样方格子分别进行统计分析[16]。从判断种群格局类型和聚集强度的角度出发,应用测定生物种群空间分布格局类型的指数,判断种群的分布格局类型[17]。在实际中,采用种群多度的方差(S2)与均值()比率(扩散系数DI)测定格局类型,然后用t检验判断分布格局的显著性,旨在避免不同指数的片面性。

式中,当DI>1时为聚集分布,当DI=1时为随机分布,当DI<1时为均匀分布。对(1)进行t检验,进一步检验种群分布格局是否偏离Poisson分布:
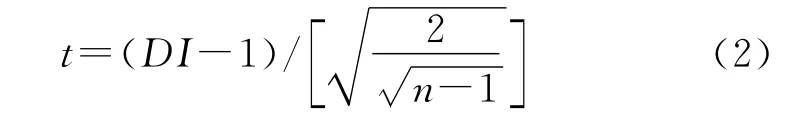
式中,n为样方数,当t>t0.05(n-1)时为聚集分布,当t≤t0.05(n-1)时为均匀分布或随机分布。
2.3 聚集度指数的测定
2.3.1 聚集度指数CI

式中,S2为样本方差,为样本均值。当CI<0时,为均匀分布;当CI=0时,为随机分布;当CI>0时,为聚集分布。
2.3.2 平均拥挤度系数M*:

式中,S2为样本方差,为样本均值。平均拥挤度是指样方内生物个体的平均邻居数,数值越大,聚集强度越大,说明一个生物个体受到其他个体的拥挤效应越大。当M*>时,为聚集分布;当M*=时,为随机分布;当M*<时,为均匀分布。
2.3.3 聚块性指数PAI

2.3.4 格林(Green)指数GI
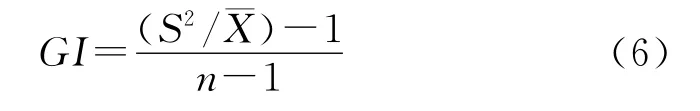
式中,S2为样本方差,为样本均值。GI>0时,为聚集分布;GI<0时,为均匀分布;GI=0时,为随机分布。
2.3.5 负二项分布指数K

式中,S2为样本方差,为样本均值。当K<0时,为均匀分布;当K>0时,为聚集分布;当K>8时,为随机分布。
2.3.6 Cassie指标(1/K)

当CA>0,为聚集分布;当CA=0时,为随机分布;当CA<0,为均匀分布。
3 结果与分析
3.1 不同海拔高度的祁连圆柏种群实际空间分布状态
根据样地调查所得祁连圆柏坐标(x,y)绘制成个体分布散点图,得到祁连圆柏种群在样地内的实际分布状态(图1)。海拔2 800m和海拔3 000m的样地中,祁连圆柏个体均较少,而海拔3 200m的样地中(样地7和8)祁连圆柏个体相对较多,并且近似集中分布于样地中间偏左或偏右。从3个海拔高度种群样地内祁连圆柏个体的实际分布状态可以看出,祁连圆柏个体在样地中数量均相对较少,说明了祁连圆柏野生种群濒危程度较高。
3.2 不同取样面积祁连圆柏种群空间分布格局类型判定
根据Poisson分布具有方差与均值相等的性质,方差/均值比率的t检验是用于检验种群分布格局是否属于随机型的一个系数,而且是建立在Poisson分布的预期假设上。由于方差反映了种群个体分布的不均匀程度,因此方差/均值比率是一个种群个体水平空间分布特征的指标[18]。利用可变尺度相邻格子样方法对祁连圆柏种群空间分布格局进行了小尺度上的理论拟合和格局强度判定。表1为在5m×5m、5m×10m和10m×10m取样面积上祁连圆柏种群的聚集强度和分布型。从表1中可以看出,在3种取样面积上,不同海拔高度祁连圆柏种群的格局指数DI>1,统计量t值远大于t0.05。因此,祁连圆柏种群空间分布格局非常显著地偏离泊松分布,表现为聚集分布。不同海拔高度祁连圆柏种群t值的大小,可以判定不同海拔梯度祁连圆柏种群分布格局皆为聚集分布,t检验进一步验证了方差/均值比的可靠性,进一步说明了聚集分布是祁连圆柏种群空间分布的基本属性。在5m×5m尺度上,祁连圆柏种群随着海拔的升高,扩散系数DI呈减小趋势;在5m×10m、10m×10m尺度上祁连圆柏种群随着海拔的升高,扩散系数DI呈先增大后减小趋势,但始终为显著聚集分布。
3.3 不同取样面积祁连圆柏种群空间分布格局的聚集强度
聚集强度是用于度量种群分布格局的聚集程度,不仅可以比较同一种群在不同时间或不同生境中聚集强度的变化,也可比较不同种群在同类生境中、同一时间的聚集状况。不同的聚集指标的测度,是从不同角度来度量同一种群聚集特性[19]。从表2可以看出,在3种取样面积上,随着取样尺度的增大,除负二项指数外,其他5个指标数值均随尺度的增大而增加,说明在所选的3个尺度中,祁连圆柏种群在10m×10m的取样尺度上分布更集中。从表2的不同海拔聚集强度看:从祁连圆柏种群分布中心的不同海拔高度,聚集强度也呈现出明显的梯度变化。祁连圆柏种群的聚集强度指数都呈减小趋势(除K外);在5m×10m、10m×10m取样面积上,祁连圆柏种群的聚集强度指数随着海拔高度的增高,都呈先增大后减小趋势(除K外),对聚集强度各指数进行排序(除K外),趋势基本一致:聚集强度大的是海拔3 000m的种群,最小的为海拔2 800m的种群,海拔3 200m的种群居中。K聚集强度的测定结果与其他几项不同,在5m×5m取样面积上,聚集强度大的是海拔3 200m的种群,聚集强度小的是海拔2 800m的种群;在5m×10m、10m×10m取样面积上,聚集强度大的是海拔3 200m的种群,聚集强度小的是海拔3 000m的种群。

图1 不同海拔高度祁连圆柏种群实际分布状态Fig.1 Spatial distribution state of S.przewalskii population in different elevation

表2 不同取样面积不同海拔梯度祁连圆柏种群聚集强度测定结果Table 2 The result of clump intensity of of S.przewalskii population in the shrub layers in different sample areas and altitudes
4 结论与讨论
种群分布格局是物种与环境之间经过长期相互适应、相互作用的结果,它与物种的生物生态学特性和种群间的竞争排斥有关,也与物种的生境有密切的联系,反映了种群个体的水平空间相互关系及其与环境条件的相互影响[20-21]。本研究结果表明,在祁连山北坡中部祁连圆柏集中分布区,祁连圆柏种群在不同取样尺度下的空间分布格局呈聚集分布,同时,在不同海拔高度下种群空间分布格局也呈聚集分布。聚集强度指标与方差/均值比率的t检验对祁连圆柏种群空间分布格局的测定结果具有良好的一致性,说明聚集分布是祁连山中段祁连圆柏自然种群空间分布格局的基本属性。这说明与前人“大多数自然种群服从集聚分布,而服从随机分布的现象较为少见[22]”的研究结果相吻合。结合野外调查的基本情况及祁连圆柏的生物学特性,我们认为祁连圆柏种群呈聚集分布的主要原因是其自身的繁殖特性和环境因子的异质性。在种子繁殖方面,祁连圆柏果实虫害严重(被害率70%左右),种子产量低,质量差(种子种仁率只有10%~20%),且后熟期长,出苗率低(一般为28%),天然更新差[23-24]。祁连圆柏由于其分布区域的局限性,主要分布在海拔2 600~4 000m的阳坡地带,属寒带气候,祁连圆柏种群呈聚集分布的分布格局,有利于增强种群的竞争力,抵御不良环境及占据与巩固营养空间。这是高海拔地区环境因子、祁连圆柏有性繁殖困难和人为干扰等多种因素综合的结果。祁连圆柏自然种群在低海拔聚集程度最低,趋向于随机分布,这与其生态学特性、人为干扰有一定关系。野外调查中还发现,低海拔分布区中放牧频繁,受人畜干扰强,在人畜干扰强的群落中,使其种子发芽及幼苗生长所需的生境条件受到破坏,在此情况下祁连山低海拔祁连圆柏种群分布格局会通过自疏机制最后趋向于随机分布。这与彭少麟、王伯荪[23]的研究结果相一致,即通过对鼎湖山森林群落种群格局的研究结果显示,一些阳生性树种均有从聚集分布向随机分布变化的趋势,阴生性树种趋向稳定顶极状态时也有聚集程度降低的倾向。
根据祁连圆柏种群的空间分布格局的动态规律,要区分阳坡、海拔的生境差异,分别采取不同的管理措施,尽量维持种群的聚集性生长。祁连圆柏种群不仅亟须有效保护,更需要采取一定的技术措施促进种群的延续与发展,要突破种群繁殖难关,扩大种群数量。由于祁连圆柏的自然生境严酷,数量稀少。种群的生存与发展难以保证,天然条件下更新繁殖率低,加强祁连圆柏繁殖技术的研究,提供一定数量的种苗以延续其种群显得非常迫切和必要。因此,本研究结果为珍稀树种祁连圆柏能够得到有效的保护提供更多的科学指导,同时也为寒带森林种群动态、群落演替的进一步研究提供理论依据。祁连圆柏种群的空间格局分析仅仅是探讨祁连山森林格局与生态过程之间关系、物种多样性维持机制的第一步,其他问题仍有待于深入研究。
[1] GREIG-SMITH P.Quantitative Plant Ecology[M].Oxford:Blackwell Scientific Publishing,1983:54-104.
[2] 张贵平,张桂萍,高昆,等.太行山南端野皂荚群落优势种群的分布格局[J].山地学报,2008,26(1):53-58.ZHANG G P,ZHANG G P,GAO K,et al.Patterns of dominant populations of Gleditsia heterophylla communities in the South of Taihang Mountains[J].Journal of Mountain Science,2008,26(1):53-58.(in Chinese)
[3] 刘富强 ,王延平 ,杨阳,等.黄河三角洲柽柳种群空间分布格局研究[J].西北林学院学报,2009,24(3):7-11.LIU F Q,WANG Y P,YANG Y,et al.Spatial distribution pattern of Tamarix chinensis in Ye1low River Delta[J].Journal of Northwest Forestry University,2009,24(3):7-11.(in Chinese)
[4] 李昌龙,马瑞君,王继和,等.甘肃民勤连古城自然保护区优势种种群结构和动态研究[J].西北植物学报,2005,25(8):1628-1636.LI C L,MA R J,WANG J H,et al.Community structures and dynamics of dominant species in Minqin Liangucheng Nature Reserve of Gansu Province[J].ActaBot.Borea1.Occident.Sin.,2005,25(8):1628-1636.(in Chinese)
[5] 何建源,卞羽,吴焰玉,等.不同坡向濒危植物南方铁杉的分布格局[J].中国农学通报,2010,26(13):122-125.HE J Y,BIAN Y,WU Y Y,et al.Spatial distribution pattern of the endangered plant Tsuga chekiangensis in different aspect[J].Chinese Agricultural Science Bulletin,2010,26(13):122-125.(in Chinese)
[6] 张金屯.数量生态学[M].北京:科学出版社,2004.
[7] 郝朝运,张小平,李文良,等.不同类型群落中濒危植物永瓣藤种群的空间分布格局[J].生态学报,2008,28(6):2900-2908.HAO C Y,ZHANG X P,LI W L,et al.Spatial distribution of Monimopetalum chinense populations in diferent forest types[J].Acta Ecologica Sinica,2008,28(6):2900-2908.(in Chinese)
[8] 蒋雪琴,刘艳红,赵本元.湖北神农架地区巴山冷杉种群结构特征与空间分布格局[J].生态学报,2009,29(5):2211-2218.JIANG X Q,LIU Y H,ZHAO B Y.Structure characteristics and spatial distribution of Abies fargesii population in Shennongjia National Nature Reserve,China [J].Acta Ecologica Sinica,2009,29(5):2211-2218.(in Chinese)
[9] WANG X C,HAN S J,ZOU C J,et al.Geostatistical analysis of the pattern of Betula ermanii population in Changbai Mountain[J].Chinese Journal of Applied Ecology,2002,13(7):781-784.
[10] ZHANG Z X,LIU P,LIU C S,et al.Studies on the population structure and spatial distribution pattern of Tsuga chinensis var.tchekiangensis[J].Jour.of Zhejiang for.Sci.&Tech.,2009,29(1):7-14.
[11] 康华靖,陈子林,刘鹏,等.大盘山自然保护区香果树种群结构与分布格局[J].生态学报,2007,27(1):389-396.KANG H J,CHEN Z L,LIU P,et al.The population structure and distribution pattern of Emmenopterys henryi in Dapanshan Natural Reserve of Zhejiang Province [J].Acta Ecologica Sinica,2007,27(1):389-396.(in Chinese)
[12] 文陇英,陈拓,张满效,等.不同生境下祁连圆柏叶片色素和稳定碳同位素组成的变化[J].冰川冻土,2010,32(4):823-828.WEN L Y,CHEN T,ZHANG M X,et al.Variations of pigments and Stable-Carbon isotope ratios in Sabina przewalskii under different environments[J].Journal of Glaciology and Geocryology,2010,32(4):823-828.(in Chinese)
[13] 苏军德,勾晓华,曹宗英,等.祁连圆柏光合作用日变化特征及其与生理生态因子的关系[J].西北植物学报,2011,31(5):1011-101.SU J D,GOU X H,CAO Z Y,et al.Diurnal course of photosynthesis and relationship with the eco-physiological factors of Qilian Juniper[J].ActaBot.Borea1.Occident.Sin.,2011,31(5):1011-1017.(in Chinese)
[14] 刘录三,邵雪梅,梁尔源,等.祁连山中部祁连圆柏生长与更新方式的树轮记录[J].地理研究,2006,25(1):53-59.LIU L S,SHAO X M,LIANG E Y,et al.Tree-ring records of Qilian Juniper's growth and regeneration patterns in the central Qilian Mountains[J].Geographical Research,2006,25(1):53-59.(in Chinese)
[15] 苏军德.祁连圆柏生理生态特性研究[D].兰州:兰州大学,2012.
[16] 刘万德,苏建荣,王梦君,等.滇西北云南红豆杉种群空间分布格局[J].西北林学院学报,2012,27(6):131-13.LIU W D,SU J R,WANG M J,et al.Spatial distribution pattern of Taxus yunnanensis population in Northwest Yunnan Province[J].Journal of Northwest Forestry University,2012,27(6):131-13.(in Chinese)
[17] 武玉珍,张峰.山西桑干河流域湿地植被优势种群分布格局研究[J].植物研究,2006,26(6):735-741.WU Y Z,ZHANG F.Patterns of dominant populations of wetland vegetation in Sanggan River Watershed,Shanxi[J].Bulletin of Botanical Research,2006,26(6):735-741.(in Chinese)
[18] 张峰,上官铁梁.山西湿地资源及可持续利用研究[J].地理研究,1999,19(4):420-427.ZHANG F,SHANGGUAN T L.A study on the wetland resources in Shanxi and their sustainale untilization[J].Geographical Research,1999,19(4):420-427.(in Chinese)
[19] 杨宁,邹冬生,李建国.衡阳盆地紫色土丘陵坡地自然恢复灌丛阶段主要种群空间分布格局[J].生态环境学报,18(3):996-1001.YANG N,ZOU D S,LI J G.Spatial pattern of main populations of the natural recovery shrub stage community in sloping-land with purple soils in Hengyang[J].Ecology and Environmental Sciences,2009,18(3):996-1001.(in Chinese)
[20] 韩路,王海珍,彭杰,等.塔里木河上游天然胡杨林种群空间分布格局与动态研究[J].西北植物学报,2007,27(8):1668-1673.HAN L,WANG H Z,PENG J,et al.Spatial distribution patterns and dynamics of major population in Populus euphraticaforest in upper reaches of Tarim River[J].ActaBot.Borea1.Occident.Sin.,2007,27(8):1668-1673.(in Chinese)
[21] 粱士楚.云贵鹅耳枥种群分布格局的分形特征[J].武汉植物学研究,2001,19(4):263-268.LIANG S C.Fractal characteristics of distribution pattern of Carpinus pubescens population[J].Journal of Wuhan Botanical Resear,2001,19(4):263-268.(in Chinese)
[22] 周纪伦.植物种群生态学[M].北京:高等教育出版社,1993,96-115.
[23] 徐生旺,蔡晋翔,仲永芳.祁连圆柏扦插育苗技术[J].陕西林业科技,2008,(1):130-131,15.
[24] 王金叶,米占山.祁连圆柏育苗技术试验研究[J].甘肃林业科技,1991,(1):22-26.
[25] 彭少麟,王伯荪.鼎湖山森林群落分析.Ⅲ种群分布格局[C]//中国科学院鼎湖山森林生态定位研究站.热带亚热带森林生态系统研究(第二集).广州:科学普及出版社,1984:24-37.
