上海地区8种落叶阔叶树夏季光合蒸腾特性研究
2015-01-02邵永昌李娟娟付达夫李二焕庄家尧
邵永昌,李娟娟,付达夫,李二焕,庄家尧*
(1.南京林业大学 水土保持与生态修复实验室,江苏 南京210037;2.迪士尼中国研究,上海200021;3.国家林业局 中南林业调查规划设计院,湖南 长沙410014)
伴随着全球气候变暖和城市化进程的加速,城市下垫面和冠层结构发生了剧烈的变化,城市热岛效应研究也成了人们关注的焦点[1-2],植被对于缓解城市热岛效应有着显著的作用[3-5],其主要是通过太阳辐射与蒸腾作用降温增湿,并通过光合作用吸收CO2并释放出O2,从而对大气湿热属性产生影响[6]。
国外学者早期对树木光合蒸腾作用的研究是以各种环境因子与光合作用之间的关系为主,试验材料主要为离体或盆栽,后来Kramer等学者对影响树木光合作用的环境因子进行了综述[7],介绍了早期树木光合生态学研究的概况;20世纪90年代以来,大气CO2浓度升高对树木光合生理的影响成了许多学者们对植物光合生理生态研究的热点,许大全[8]简要评述了树木光合作用对CO2浓度升高的响应,以及影响这种响应的因素及光合适应的可能机理。蒋高明[9]等对国外近几十年在CO2浓度升高对植物的直接影响方面所展开的生理生态学研究方法、动态、基本理论和存在问题等内容作了介绍;对于城市植物的光合生理研究多为园林绿化植物,如金叶假连翘、花叶鹅掌藤、金边决明、狗牙花、北美枫香、美国红栌等[10-12];城市绿化树种对外界环境改变的响应机制的本质和如何实现由叶片光合到整株植物的尺度转是城市绿化树种光合生理生态研究中亟待解决的问题[13];早期学者们对树木蒸腾耗水的研究主要采用快速称重的方法,20世纪90年代以来,树木耗水特性的测定方法主要以热脉冲法、热平衡法和热扩散法、气孔计法、和微气象法为主,稳定性示踪物(氘)也得到了相应的发展[14];虽然对树木蒸腾耗水的方法研究已趋于完善,但是测定结果需用传统方法的测定结果加以校正,因此,找到一个相对标准的校正方法是今后树木研究的主要方向[15]。
光合作用和蒸腾作用是植物重要的生命活动,光合作用是植物生长的生理基础,蒸腾作用耗水则是树木水分散失的主要途径,而水分利用效率则是评价植物水分消耗和利用能力的主要评价指标[16]。因此,探讨常见绿化树种的光合作用特性,主要环境因子对绿化植物光合、蒸腾作用的影响,确定主要绿化树种光合因子对植物光合、蒸腾作用的影响,对植物叶片水分利用效率和光能利用效率的影响是十分必要的,同时也为城市环境绿化和环境保护提供相关研究理论依据。在对上海市现有的植物种类和生长状况进行调查与分析的基础上,根据树种的重要值与出现的频率等综合指标,选择具有代表性的8个阔叶落叶树种,拟从树种的光合作用特征角度出发,对上海地区常见8种落叶阔叶树种的光合蒸腾特性日变化、水分利用效率、光能利用效率和环境因子的相关性分析等进行综合分析比较,探讨其光合蒸腾特性和水分生理生态特性,为城市森林的人工植被群落配置和改造、建筑周边环境绿化、城市行道树种选择提供参考。
1 材料与方法
1.1 试验地概况
研究区位于上海市浦东川沙园林苗圃基地(121°39′E,31°08′N),属北亚热带季风性气候,雨热同期,日照充分,雨量充沛,年均降水量1 048~1 138mm。上海市夏季酷热,冬季湿冷,年均温15.2~15.9℃,极端最高气温40.8℃(2013年8月7日),极端最低气温-2.1℃(1893年1月19日),春秋较短,冬夏较长。
1.2 试验材料
试验材料为上海浦东川沙苗圃基地内8个阔叶落叶树种的12~15年生大苗,分别为栾树(Koelreuteria paniculata)、白 玉 兰 (Magnolia denudata)、无患子(Sapindus mukorossi)、榉树(Zelkova schneideriana)、鹅掌楸(Liriodendron chinense)、银杏(Ginkgo biloba)、悬铃木(Platanus)、三角枫(A-cer buergerianum),均为上海地区常见绿化树种,这些树种均配备地面灌溉措施,土壤水分能够得到及时补充,植株均长势良好(表1)。

表1 树种基本情况统计Table 1 Tree statistics of the basic situations
1.3 研究方法
于2013年7月中旬(晴天),使用Li-6400XT便携式光合作用测定系统,采用标准叶室选取各树种中部冠层阳面中高部位树枝前端第3~5片成熟功能叶片,从7:30-17:30每2h测定净光合速率(Pn,μmol·m-2·s-1)、蒸腾速率(Tr,mmol·m-2·s-1)、气孔导度(Gs,mol·m-2·s-1)、胞间 CO2浓度(Ci,μmol·mol-1)、叶片温度(T1,℃)及主要环境因子水汽压亏缺(VPD,kPa)、大气CO2浓度(Ca,μmol·mol-1)、光合有效辐射(PAR,μmol·m-2·s-1)、气温(Ta,℃)、相对湿度(RH,%)等参数,重复测定3株·树种-1,取平均值。根据测定的相关参数计算得出下列相关指标[17]:气孔限制值Ls=1-Ci/Ca,水分利用效率 WUE(μmol·mmol-1)=Pn/Tr;光能利用效率LUE/%=Pn/PAR×100
1.4 数据分析
使用 Microsoft Excel 2007软件进行数据处理和表格制作,Origin 8.5绘制图件,采用SPSS 19.0软件进行相关的统计与分析。
2 结果与分析
2.1 主要环境因子的日变化
观测时间内(7:30-17:30)光合有效辐射(PAR)呈单峰曲线变化,13:30左右达到最大值(1 790μmol·m-2·s-1),随后逐渐减弱;经过夜间植物呼吸作用的富集,大气CO2浓度(Ca)在7:30表现为最高值(365μmol·mol-1),在15:30左右形成一个低谷(349μmol·mol-1),此后有所回升,呈先下降后上升的趋势,与PAR日变化规律相反,却有滞后(图1A);随着PAR的逐渐增大,气温(Ta)亦逐步升高,呈单峰型曲线,在11:30-15:30空气温度较高,最高温度出现在14:30(34.85℃),相对于PAR变化有所滞后,日变化幅度较大,温差达7℃;空气相对湿度(RH)日变化与气温相反,呈“U”形曲线变化,7:30时RH 最高(70.45%),随着PAR和Ta的升高,RH 逐渐降低,在13:30达到全天最低值(51.95%),之后开始回升(图1B);RH 与PAR、Ta分别呈显著(p<0.05)、极显著(p<0.01)的负相关。
2.2 净光合速率(Pn)日变化
由图1和图2可知,8个树种净光合速率(Pn)日变化曲线在测定时段均为双峰型,上午随着气温(Ta)和光合有效辐射(PAR)的增大,Pn随之升高,之后随着温度和有效辐射的继续增大而降低,下午光强逐渐减弱,Pn有一定程度的增大,最后随着PAR和Ta的降低,Pn随之减小,三角枫第1个峰值出现在9:30,其他树种(无患子、白玉兰、鹅掌楸、栾树、银杏、榉树、悬铃木)第1个峰值出现在11:30,第2个峰值均出现在15:30,都具有明显的“光合午休”现象,可能是由于午间强烈的太阳辐射使气温升高,相对湿度湿度降低,蒸腾强烈,植物体内水分不足引起气孔关闭导致Pn下降[18],Pn午间降低的程度不同说明8个树种受环境因子的抑制各不相同,不同树种Pn的峰值发生时间的不同又说明8个树种的光合作用对自然环境因子的响应具有一定的差异性;无患子、白玉兰、鹅掌楸具有较高的日均净光合速率,栾树日均净光合速率较低(表2)。
2.3 蒸腾速率(Tr)和气孔导度(Gs)日变化
悬铃木、三角枫、栾树、白玉兰、榉树、鹅掌楸、银杏蒸腾速率(Tr)日变化均为双峰曲线,悬铃木、榉树、鹅掌楸、银杏第1个峰值出现在11:30,三角枫、白玉兰、栾树第1个峰值出现在9:30,7个树种第2个峰值均出现在15:30,无患子Tr日变化为单峰曲线,最大值出现在11:30(3.32mmol·m-2·s-1);就蒸腾速率日变化而言,悬铃木、三角枫、无患子、银杏、白玉兰表现出中午前后(9:30-15:30)较高,早上和傍晚较低,鹅掌楸、榉树午间(11:30-13:30)较高,榉树和栾树在午间出现蒸腾速率的轻微下降可能是由于中午温度高,失水过多,导致气孔部分关闭所致[19](图3A),其他树种在中午前后Tr维持一个较高的水平,因为植物会通过提高蒸腾速率以降低叶温来维持叶片各器官的正常功能;蒸腾速率在一定程度上反映了植物适应逆境调节水分损失的能力和适应干旱环境的能力[20],白玉兰、无患子、榉树、三角枫具有较高的蒸腾速率,栾树蒸腾速率较低(表2),相同环境下,白玉兰、无患子、榉树、三角枫较其他树种单位叶面积要消耗更多的水分,也意味着这4个树种叶片单位面积失水量的大幅增加,只有及时保证植株水分的充足供应才能保证植物叶片的水分供给。
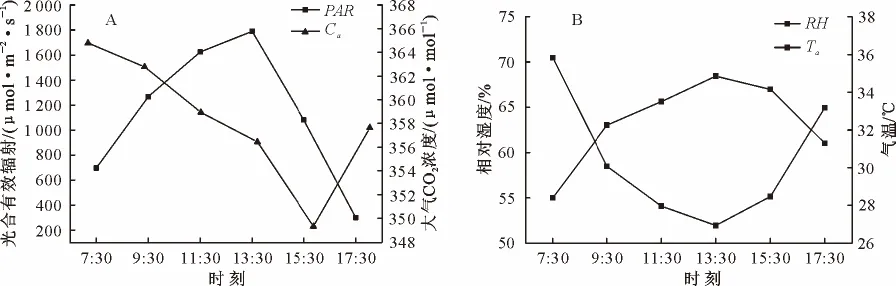
图1 主要环境因子日变化Fig.1 Diurnal changes of the main environmental factors

图2 不同树种净光合速率(Pn)日变化曲线Fig.2 Diurnal changes of net photosynthetic rate of different species
8个树种气孔导度(Gs)日变化曲线在测定时段均为双峰型,Gs与其对应的蒸腾速率(Tr)日变化规律基本相似,早上可能由于气温较低,导致CO2扩散能力降低,叶片的Gs值相对较小,上午随着光强的增大,气孔缓慢张开[21],Gs随之上升,在Pn和Tr增大到一定程度后Gs出现不同程度的下降;除三角枫外,其他树种(无患子、悬铃木、银杏、榉树、鹅掌楸)峰值出现的时间和其对应的Tr峰值出现的时间相同(图3A)。

图3 不同树种蒸腾速率(Tr)和气孔导度(Gs)日变化曲线Fig.3 Diurnal changes of net photosynthetic rate and stomatal conductance of different species
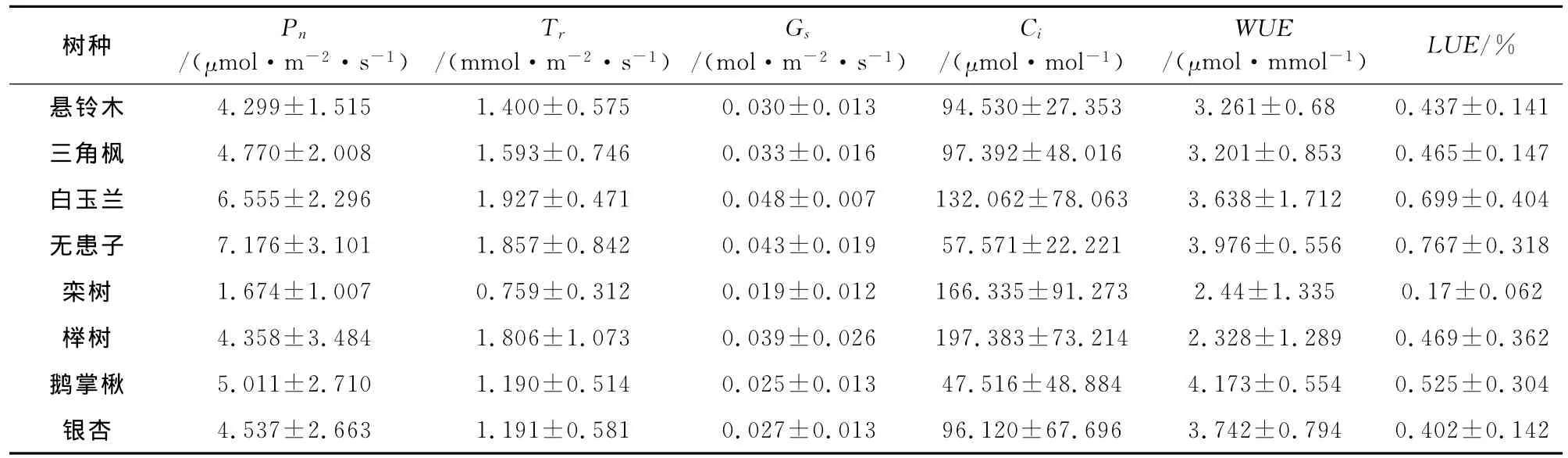
表2 不同树种光合参数日平均值Table 2 Daily average photosynthetic parameters of different species
2.4 胞间CO2浓度(Ci)日变化和气孔限制值(Ls)日变化
胞间CO2浓度(Ci)的变化直接影响植物提供光合作用碳源的多少,Ci下降,则表明植物光合作用速率大于呼吸速率,说明植物同化利用CO2[30],反之,Ci升高,说明植物光合作用速率降低;由图1、2和4A可知,8个树种胞间CO2浓度(Ci)日变化在测定时段呈“W”形,上午Ci随着净光合速率(Pn)的升高而逐渐降低,在光强和气温相对较高的午间前后,8个树种Ci都有不同程度的升高,随着植物“光合午休”现象的消失,Ci也随之降低,15:30之后光强和气温下降,叶片光合作用减弱,Ci开始升高;无患子、悬铃木、三角枫第一个谷值出现在9:30,其他树种(鹅掌楸、白玉兰、榉树、银杏、栾树)第1个谷值出现在11:30,第2个谷值均出现在15:30(图4A),叶片Pn日变化过程与Ci之间呈负相关(表3),这种负相关表明Pn随着光强的上升而增大的主要原因是叶肉细胞的光合活性的增大,从而也导致了Ci的减小。
通常认为判断叶片净光合速率(Pn)降低的原因是根据胞间CO2浓度(Ci)和气孔限制值(Ls)的变化趋势,即当Ls增大、Ci降低时认为Pn下降原因是Gs降低,为气孔限制;Ls减小、Ci增大或不变时认为Pn下降的原因为非气孔限制[22-23]。根据图2、3A、4可以得出,榉树、栾树、白玉兰、鹅掌楸在11:30-13:30和三角枫在9:30-11:30的Pn下降为气孔限制,无患子、三角枫、悬铃木、银杏在11:30-13:30的Pn下降为非气孔限制,则表明树种在午间前后Pn下降的因素表现出一定的差异性,但15:30之后Pn下降均是因为非气孔限制,说明在一定光强和气温下,无患子、三角枫、悬铃木、银杏Pn下降是由于光合组织的功能受到了损害而导致的。

图4 不同树种胞间CO2(Ci)浓度和气孔限制值(Ls)日变化Fig.4 Diurnal changes of stomatal CO2concentration and stomatal limiting value of different species
2.5 水分利用效率(WUE)和光能利用效率(LUE)日变化
植物水分利用效率(WUE)是指植物消耗单位水分所生产的同化物质的量,包括单株整体水平和单叶生理水平等方式表示,它实质上反映了植物耗水与其干物质生产之间的关系,也是反映植物耗水和耐旱特性的指标之一[24]。由图5A可知,榉树、栾树、白玉兰WUE日变化波动较大,其他树种较为平缓,表明榉树、栾树WUE对环境因子的响应比其他树种更为敏感,鹅掌楸、银杏、三角枫WUE日变化均表现出上午高于下午并随时间逐渐降低的特点;就日平均值而言,鹅掌楸、无患子、银杏、白玉兰具有较高的WUE,分别为4.173、3.976、3.742μmol·mmol-1和3.638μmol·mmol-1,而三角枫、悬铃木、栾树、榉树则相对较低,分别为3.261、3.201、2.440μmol·mmol-1和2.328μmol·mmol-1(表2),表明前4个树种比后4个树种能够更有效地利用土壤水分,更易在干旱条件下存活。
光能利用效率反映了植物对太阳能的利用能力,通过植物叶片光合组织固定的碳量表现出来[25]。由图1、5B可知,8个树种LUE日变化曲线均在午后(13:30左右)出现最低值,这与中午较强的太阳辐射和树种的“光合午休”现象有关;下午15:30之后,由于光合有效辐射(PAR)的迅速下降,树种的LUE都出现上升的趋势,17:30达到最大值;经相关性分析,植物LUE和净光合速率(Pn)、光合有效辐射(PAR)分别呈极显著正相关(p<0.01)、显著负相关(p<0.05)的关系(表3),说明光强达到一定程度后,植物对太阳能的利用效率随着光强的升高而降低;就日均值而言,无患子、白玉兰和鹅掌楸具有较高的LUE,分别为0.767%、0.699%、0.525%,而榉树、三角枫、悬铃木、银杏、栾树相对较低,分别为0.469%、0.465%、0.437%、0.402%、0.170%(表2),表明前3个树种在相同的光合有效辐射下,单位叶面积能够固定更多的碳量,对太阳能具有更强的利用能力。
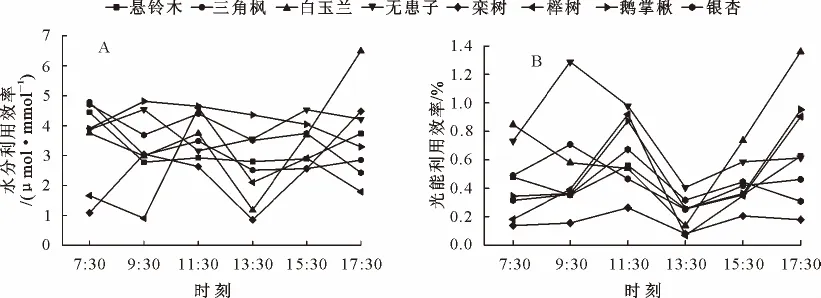
图5 不同树种水分利用效率(WUE)和光能利用效率(LUE)日变化曲线Fig.5 Diurnal changes of water use efficiency and light use efficiency of different species
2.6 环境因子相关性分析
2.6.1 净光合速率(Pn)和环境因子的相关性分析
如表3所示,除白玉兰外,其他7个树种Pn与PAR、Ta呈正相关,测试树种均与RH、Ca呈负相关,一定程度上,气温和太阳辐射越高越有利于植物光合作用的进行;除三角枫、白玉兰外,6个树种Pn与Vpd、T1均为正相关,但相关性不明显(表3)。

表3 不同树种净光合速率(Pn)与光合参数相关性Table 3 Correlation analysis on photosynthetic parameters of different species
2.6.2 蒸腾速率(Tr)与环境因子相关性分析 由表4可知,8个树种Tr与PAR、Ta、Vpd、T1呈正相关,Tr与RH、Ca均为负相关,空气相对湿度越大、大气CO2浓度越高则植物蒸腾速率越小。
3 结论与讨论
8个树种净光合速率(Pn)日变化在测定时段均为双峰型,存在明显的“光合午休”现象,许多学者试图从同化物积累、生理节奏等角度对其“午休”机理进行解释,但迄今尚无定论[26];8个树种净光合速率峰值出现时间和午间降低程度的不同,说明其受环境因子的抑制各不相同,不同树种光合作用对环境因子的响应具有一定的差异性,无患子、白玉兰、鹅掌楸具有较高的日均净光合速率,分别为7.176、6.555μmol·m-2·s-1和5.011μmol·m-2·s-1,说明无患子的光合能力和光合潜力较强,适用于阳性区域大面积种植,栾树日均净光合速率较低,为1.674μmol·m-2·s-1,说明其耐受能力较差。

表4 不同树种蒸腾速率(Tr)与光合参数相关性Table 4 Correlation analysis on transpiration rate of different species
榉树、栾树、白玉兰、鹅掌楸午间Pn的下降为气孔限制,无患子、三角枫、悬铃木、银杏在午间的Pn下降为非气孔限制,说明在一定光强和气温下,无患子、三角枫、悬铃木、银杏Pn下降是由于光合组织的功能受到了损害而导致,而榉树、栾树、白玉兰、鹅掌楸是由于植株蒸腾作用强烈、叶片失水过多而导致气孔导度(Gs)降低引起的[23],因此,适当控制环境因子则是提高植物光合产量的重要措施,如通过遮光措施来控制光照强度,适当的灌溉措施来提高植物水分的补给和周边环境空气的湿度等。
悬铃木、三角枫、栾树、白玉兰、榉树、鹅掌楸、银杏蒸腾速率(Tr)日变化均为双峰曲线,无患子Tr日变化为单峰曲线;蒸腾速率决定着植物体内水分的平衡,也在一定程度上反映出植物适应干旱环境并调节水分损失的特性[27]。相同环境条的件下,白玉兰、无患子、榉树、三角枫单位叶面积要消耗更多的水分,其通过蒸腾作用产生降温增湿效果对周边小气候环境具有更强的调节能力,但同时需要注意在夏季高温期间的园林绿化实践中应结合植物的冠层结构等因素进行合理的灌溉管理,以保证植株有充足的水分供应。
8个树种气孔导度(Gs)日变化曲线在测定时段均为双峰型,与其对应的蒸腾速率日变化规律相似,这与何春霞等的研究结果一致[19],已有研究将土壤水分纳入到了气孔导度的模拟过程中。本次研究并没有考虑土壤水分对蒸腾速率和气孔导度的影响,植物叶片蒸腾速率与土壤水分条件关系密切,特别是在干旱半干旱条件下[29],在今后的研究中需要进一步完善[28]。
植物水分利用效率(WUE)反映了植物的耗水和耐旱特性,榉树、栾树的WUE对环境因子的响应较其他树种更为敏感,鹅掌楸、无患子、银杏、白玉兰具有更高的水分利用效率,分别为4.173、3.976、3.742μmol·mmol-1和3.638μmol·mmol-1,表明这4个树种能够更有效地利用土壤水分,在极端环境条件下具有更好的适应性,分析植物水分利用效率还需要考虑植物的灌溉状况,本次研究树种均具有良好的灌溉条件,干旱半干旱条件下,植物的水分利用效率必将受到土壤水分供给的影响,还需要进一步研究。
8个树种光能利用效率日变化均在午后出现最低值,这与中午较强的太阳辐射和树种的“光合午休”现象有关;光强达到一定程度后,植物对太阳能的利用效率随着光强的升高而降低,无患子、白玉兰和鹅掌楸具有较高的LUE日均值,分别为0.767%、0.699%、0.525%,在相同的环境条件下,无患子、白玉兰、鹅掌楸单位叶面积能够固定更多的碳量,对太阳能具有更强的利用能力。
由于本次研究是在一定的灌溉条件下进行的,对比其他学者的研究分析,本次试验中,悬铃木、三角枫、白玉兰、无患子、榉树、鹅掌楸的净光合速率分别较非灌溉条件下提高59%、19%、21%、24%、192%、163%[19,31-33],蒸腾速率分别较非灌溉条件下高出67%、34%、35%、18%、64%、121%[19,31-33],水分利用效率分别较非灌溉条件下低出3%、6%、17%、29%、14%、68%[19,31-33],相较非灌溉条件,灌溉条件下树种表现为高光合、高蒸腾、低水分利用效率的特点,这可能是由于植株水源充盈,蒸腾速率大大提高,一定程度上降低了植株的水分利用效率。
不同树种净光合速率(Pn)、蒸腾速率(Tr)与环境因子的相关性分析表明,一定条件下,光强、气温越高越有利于气孔的张开,则叶片净光合速率和蒸腾速率就越大,但是树种有光饱和点且光合作用有“午休”现象[26],这与环境因子和树种的内部生理因素有关,由于榉树、栾树、鹅掌楸在11:30-13:30的Pn下降为气孔限制,说明这种Pn与Ci的负相关是由于叶片光合组织活性的变化导致的[22];叶片是植物与外界环境之间气体和水分交换的主要通道和调节器官,是SPAC系统物质与能量交换的重要调控通道,8个树种Tr与RH、Ca均为负相关关系,说明在一定条件下空气相对湿度越大、大气CO2浓度越高则植物蒸腾速率越小。
致谢:感谢迪士尼中国研究对本研究课题提供资金和学术观点贡献。
[1] UNHABITAT.State of the world's cities 2010/2011-bridg-ing the urban divide[M].London:Earthscan,2008:244.
[2] OKE T R.Boundary layer climate[M].Cambridge:Great Britain at the University Press,1987:1-3.
[3] FAUNCE T,STYRING S,WASIELEWSKI M R,et al.Artificial photosynthesis as a frontier technology for energy sustainability[J].Energy &Environmental Science,2013,6(4):1074-1076.
[4] LI Q,ZHANG H,LIU X,et al.Urban heat island effect on annualmean temperature during the last 50years in China[J].Theoretical and Applied Climatology,2004,79(3):165-174.
[5] GAGO J,COOPMAN R E,CABRERA H M,et al.Photosynthesis limitations in three fern species[J].Physiologia Plantarum,2013,149(4):599-611.
[6] GATES D M.Transpiration and leaf temperature[J].Ann.Review Plant Physiology,1968,19:211-238.
[7] 张赐成.桂花树光合作用特征分析[D].长沙:湖南师范大学,2014.
[8] 许大全.光合作用及有关过程对长期高CO2浓度的响应[J].植物生理学通讯,1994(2):81-87.XU D Q.Responses of photosynthesis and related processes to long-term high CO2concentration[J].Plant Physiology Communications,1994(2):81-87.(in Chinese)
[9] 蒋高明,韩兴国,林光辉.大气CO2浓度升高对植物的直接影响——国外十余年来模拟实验研究之主要手段及基本结论[J].植物生态学报,1997(6):2-15.JIANG G M,HAN X G,LIN G H.Response of plant growth to elevated[CO2]:a review on the chief methods and basic conclusions based on experiments in the external countries in past decade[J].Acta Phytoecologica Sinica,1997(6):2-15.(in Chinese)
[10] 俞继红.3种彩叶植物的光合特性比较[J].西北林学院学报,2014,29(4):21-25 YU J H.A comparison of photosynthetic characteristics between three color-leafed plants[J].Journal of Northwest Forestry University,2014,29(4):21-25.(in Chinese)
[11] 杭夏子,翁殊斐,袁喆.华南5种园林灌木叶性状特征及其对环境响应的研究[J].西北林学院学报,2014,29(2):243-247.HANG X Z,WENG S F,YUAN Z.Relationships between leaf traits of 5plantscape shrubs and their responses to the environment in Southern China[J].Journal of Northwest Forestry University,2014,29(2):243-247.(in Chinese)
[12] 吴志庄,李伟成,熊德礼,等.不同竹龄青皮竹光合作用与荧光特性的比较[J].西北林学院学报,2013,28(6):33-36.WU Z Z,LI W C,XIONG D L,et al.A comparative study on photosynthetic and fluorescence characteristicsamong different aged bambusa textilis[J].Journal of Northwest Forestry University,2013,28(6):33-36.(in Chinese)
[13] 陈德祥,李意德,骆土寿,等.热带森林树种光合作用生理生态研究进展[J].江西农业大学学报,2004(6):905-911.CHEN D X,LI Y D,LUO T S,et al.Progress in physioecology research of photosynthesis in tropical forest[J].Acta Agriculturae Universitatis Jiangxiensis,2004(6):905-911.(in Chinese)
[14] 孔俊杰,贾黎明,李广德.影响树木蒸腾耗水的外部因子研究进展[J].世界林业研究,2007(1):16-21.KONG J J,JIA L M,LI G D.Advances in the researches of exterior effect factors on tree water consumption[J].World Forestry Research,2007(1):16-21.(in Chinese)
[15] 郭孟霞,毕华兴,刘鑫,等.树木蒸腾耗水研究进展[J].中国水土保持科学,2006(4):114-120.GUO M X,BI H X,LIU X,et al.Review on the water consumption of tree transpiration[J].Science of Soil and Water Conservation,2006(4):114-120.(in Chinese)
[16] 张艳丽,费世民,李智勇,等.成都市沙河主要绿化树种固碳释氧和降温增湿效益[J].生态学报,2013,33(12):3878-3887.ZHANG Y L,FEI S M,LI Z Y,et al.Carbon sequestration and oxygen release as well as cooling and humidification efficiency of the main greening tree species of Sha River,Chengdu[J].Acta Ecologica Sinica,2013,33(12):3878-3887.(in Chinese)
[17] SAGE R F,SAGE T L,Kocacinar F.Photorespiration and the evolution of C4photosynthesis[J].Annual Review of Plant Biology,2012,63:19-47.
[18] 康雅茸,张华.兰州银滩黄河湿地不同植物夏季光合日变化[J].生态学杂志,2014,33(9):2336-2342.KANG Y R,ZHANG H.Diurnal variation of photosynthesis of different plants in summer in Yintan wetland of Lanzhou section of Yellow River[J].Chinese Journal of Ecology,2014,33(9):2336-2342.(in Chinese)
[19] 何春霞,李吉跃,郭明,等.4种乔木叶片光合特性和水分利用效率随树高的变化[J].生态学报,2008,28(7):3008-3016.HE C X,LI J Y,GUO M,et al.Changes of leaf photosynthetic characteristics and water use efficiency along tree height of 4tree species[J].Acta Ecologica Sinica,2008,28(7):3008-3016.(in Chinese)
[20] 田晶会,贺康宁,王百田,等.黄土半干旱区侧柏蒸腾作用及其与环境因子的关系[J].北京林业大学学报,2005(3):53-56.TIAN J H,HE K N,WANG B T,et al.Relationship between transpiration of Platycladus orientalisandenvironmental factors in semi_arid region on Loess Plateau[J].Journal of Beijing Forestry University,2005(3):53-56.(in Chinese)
[21] 邵玺文,韩梅,韩忠明,等.不同生境条件下黄芩光合日变化与环境因子的关系[J].生态学报,2009,29(3):1470-1477.SHAO X W,HAN M,HAN Z M,et al.Relationship between diurnal changes of photosynthesis of Scutellaria baicalensis and environmental factors in different habitats[J].Acta Ecologica Sinica,2009,29(3):1470-1477.(in Chinese)
[22] FARQUHAR G D,SHARKEY T D.Stomatal conductance and photosynthesis[J].Annual Review of Plant Physiology,1982,33:317-345.
[23] 许大全.光合作用气孔限制分析中的一些问题[J].植物生理学通讯,1997(4):241-244.XU D Q.Some problems in stomatal limitation analysis of photosynthesis[J].Plant Physiology Communications,1997(4):241-244.(in Chinese)
[24] 宋锋惠,罗青红,史彦江,等.新疆杂交榛良种光合作用对生态因子的响应[J].生态学杂志,2013,32(5):1148-1154.SONG F H,LUO Q H,SHI Y J,et al.Photosynthetic responses of hybrid hazel fine breeds to ecological factors in Xinjiang,Northwest China[J].Chinese Journal of Ecology,2013,32(5):1148-1154.(in Chinese)
[25] 何东进,洪伟,吴承祯,等.毛竹种群光能利用率的研究[J].福建林学院学报,1999(4):324-326.HE D J,HONG W,WU C Z,et al.Study on light energy utilization percent of Phyllostachys pubescens population[J].Ournal of Fujian College of Forestry.1999(4):324-326.(in Chinese)
[26] 许大全.光合作用的“午睡”现象[J].植物生理学通讯,1997(6):466-467.
[27] 刘明虎,辛智鸣,徐军,等.干旱区植物叶片大小对叶表面蒸腾及叶温的影响[J].植物生态学报,2013(5):436-442.LIU M H,XIN Z M,XU J,et al.Influence of leaf size of plant on leaf transpiration and temperature in arid regionsp[J].Chinese Journal of Plant Ecology,2013(5):436-442.(in Chinese)
[28] GAO Q,ZHAO P,ZENG X,et al.Amodelofstomatal conductance to quantify the relationship between leaf transpiration,microclimate and soilwater stress[J].Plant,Cell and Environment,2002,25:1373-1381.
[29] 张华,王百田,郑培龙.黄土半干旱区不同土壤水分条件下刺槐蒸腾速率的研究[J].水土保持学报,2006(2):122-125.ZHANG H,WANG B T,ZHENG P L.Study on transpiration rate of Black Locust under different soil water content in Semi-Arid region Loess Plateau[J].Journal of Soil and Water Conservation,2006(2):122-125.(in Chinese)
[30] 赵顺,黄秋娴,李玉灵,等.遮荫处理对臭柏幼苗光合特性的影响[J].生态学报,2014,34(8):1994-2002.ZHAO S,HUANG Q X,LI Y L,et al.Effects of shading treatments on photosynthetic characteristics of Juniperus sabina Ant.seedlings[J].Acta Ecologica Sinica,2014,34(8):1994-2002.(in Chinese)
[31] 梁淑英,胡海波,夏尚光.枫杨、悬铃木和女贞光合特性的比较[J].南京林业大学学报:自然科学版,2008(2):135-138.LIANG S Y,HU H B,XIA S G.Comparison of photosynthetic characteristics among Pterocarya stenoptera,Platanus acerifolia and Ligustrum lucidum [J].Journal of Nanjing Forestry University:Natural Sciences Edition,2008(2):135-138.(in Chinese)
[32] 张娇,施拥军,朱月清,等.浙北地区常见绿化树种光合固碳特征[J].生态学报,2013,33(6):1740-1750.ZHANG J,SHI Y J,ZHU Y Q,et al.The photosynthetic carbon fixation characteristics of common tree species in Northern Zhejiang[J].Acta Ecologica Sinica,2013,33(6):1740-1750.(in Chinese)
[33] 程建峰,姜锟,沈允钢.修枝截干对二球悬铃木叶片光合特性的影响[J].中国生态农业学报,2009,17(3):469-473.CHENG J F,JIANG K,SHEN Y G.Effect of branch and stem pruning on photosynthetic characteristics of Platanus acerifolia L.[J].Chinese Journal of Eco-Agriculture,2009,17(3):469-473.(in Chinese)
