铁、钙离子补料对杜氏盐藻生长的影响
2014-02-23吕和鑫贾士儒崔相敢夏锋
吕和鑫,贾士儒,崔相敢,夏锋
(天津科技大学天津市工业微生物重点实验室,天津300457)
铁、钙离子补料对杜氏盐藻生长的影响
吕和鑫,贾士儒*,崔相敢,夏锋
(天津科技大学天津市工业微生物重点实验室,天津300457)
通过原子吸收光谱测定了杜氏盐藻对钙、镁、钾、铁四种离子的消耗速率,构建金属离子含量限制型培养基,研究了补加金属离子对杜氏盐藻生长光合速率、色素含量的影响。结果表明,对数期杜氏盐藻细胞对铁离子与钙离子的消耗呈线性降低,每增殖106个细胞分别消耗0.042 μg的铁离子与0.708 μg的钙离子,而在稳定期后,钙、铁离子浓度继续下降。生长时期中钾离子与镁离子的含量变化不显著。铁、钙离子补料能恢复杜氏盐藻的生长,但光合活性与色素含量变化较补料前的对数期光合速率与色素含量要低,但总类胡萝卜素含量要高于补料前对数期的。培养到稳定期的培养基经过回收补加有限的铁、钙离子后,细胞生长延滞期消失,但稳定期细胞浓度降低。回流培养基对数期细胞的光合速率与叶绿素含量降低,但是总类胡萝卜素含量升高。
杜氏盐藻;β-胡萝卜素;金属离子;消耗速率;基质消耗动力学
杜氏盐藻(Dunaliella salina)是一种在胁迫生长条件下可大量产生具有抗氧化活性的β-胡萝卜素(β-carotene)与黄体素(lutein)的单细胞海洋绿藻,其产生的β-胡萝卜素被用作食品添加剂与着色剂、饲料添加剂、复合维生素制剂、化妆品中的抗氧化剂等[1]。杜氏盐藻在生长环境适宜的条件下具有一对鞭毛,使细胞具有游动能力,但是在一些如营养缺乏、低温、高光照、高盐度等极端环境条件下,盐藻细胞会发生一些如鞭毛脱落等形态与油脂积累、胡萝卜素积累等生化方面的调整以便更好地适应环境变化[1-2]。胡萝卜素可以捕获光能防止光合系统光氧化损伤的发生[3-5],可以增强叶绿体膜在氧化胁迫状态下的稳定性[1,6]。杜氏盐藻中胡萝卜素主要积累于叶绿体中的内囊体之间的油质体(oil globules)中,使得细胞呈现橙色或红色。有报道表明,杜氏盐藻细胞中胡萝卜素的积累量与一个细胞分裂周期中接收的光照多少成正比,胡萝卜素的积累量受到高光照辐射、高盐度、温度胁迫以及营养胁迫的促进[2]。COESEL S N等[7]的实验表明,杜氏盐藻在营养充足时,单独或同时施加高光照与高盐度胁迫均不能诱导胡萝卜素的积累。而在营养胁迫的培养条件下,单独或同时施加高光照与高盐度均能促进胡萝卜素的积累,稳定状态的八氢番茄红素合成酶(phytoene synthase)与八氢番茄红素去饱和酶(phytoene desaturase)的信使核糖核酸(messenger ribonucleic acid,mRNA)的表达量也同时增高。所以,营养胁迫是杜氏盐藻胡萝卜素合成的前提条件,而高光照与高盐度可以促进胡萝卜素的积累速度与水平。
因此,研究盐藻培养中各矿质营养盐的动态消耗情况对于理解胡萝卜素合成途径与矿质营养盐在调控上的关系以及实现盐藻大规模生产中控制胡萝卜素合成的时间与含量具有较大的理论与应用价值。金属离子是细胞必需的营养成分,可作为酶的活性结构与调节成分、参与一些活性分子如色素的结构维持、调节细胞内的离子与渗透压平衡等。本研究通过跟踪盐藻分批发酵培养中的对数期与稳定期钾钙镁铁金属离子的浓度,确定了盐藻培养过程中铁、钙、镁与钾四种金属离子的消耗速率。
1 材料与方法
1.1 材料与试剂
杜氏盐藻(Dunaliella salina):天津科技大学天津市工业微生物重点实验室保存。
采用改良的Johnsons培养基[8]:MgCl2·7H2O 1.5 g/L,MgSO4·7H2O 0.5 g/L,KNO30.5 g/L,KCl 0.2 g/L,NaHCO30.151 g/L,KH2PO40.043 g/L,CaCl20.035 g/L,NaCl 30.0 g/L,Na2EDTA·2H2O 2.09 mg/L,FeCl3·6H2O 2.44 mg/L,H3BO30.61mg/L,(NH4)6Mo7O24·4H2O0.38mg/L,ZnCl20.041mg/L,CuSO4·5H2O 0.06 mg/L,CoCl2·6H2O 0.051 mg/L,MnCl2· 4H2O 0.041 mg/L。
其他试剂均为分析纯:天津化学试剂三厂。
1.2 仪器与设备
岛津AA-6800原子吸收光谱仪、岛津UVmini-1240紫外可见分光光度计:日本岛津有限公司;PHSJ-4A pH计:上海雷磁仪器厂;LD5-2A台式高速离心机:上海医用分析仪器厂。
1.3 试验方法
1.3.1 培养方法
利用250 mL玻璃摇瓶,加入改良的Johnsons培养基[8]静置培养,培养体系均是150 mL。每天轻轻摇动若干次使藻液混匀。培养光照强度为60 μmol/(m2·s),培养温度保持为27℃。每次接种均为对数期细胞,接种光密度(OD430nm)为0.1,接种后给予两个连续的光暗周期(12 h/12 h)以同步化细胞的生长时期,两个光周期后采用连续光照培养。
铁、钙离子限制型培养基的配制:铁离子为0.123 mg/L;钙离子为2.07 mg/L;其他离子量同Johnsons培养基,细胞密度0.15×106CFU/mL。离心收集的藻细胞用新鲜铁、钙离子限制型培养基漂洗3次后接种,接种光密度(OD430nm)为0.1。
铁、钙回流培养基的配制:离心去除稳定期藻细胞后的培养基利用0.22 μm滤膜过滤后,回流培养基的中的铁、钙离子的质量浓度为35.31 μg/L与4.86 μg/mL,补加铁离子质量浓度至0.187 mg/L,其他离子量同Johnsons培养基。藻细胞用回流培养基漂洗3次后接种。接种光密度(OD430nm)为0.1,接种细胞密度为0.15×106CFU/mL。
2.印尼北干巴鲁领队连俊伟先生之感言:(我们)虽然回到家,心却留在长沙,每天几乎还紧盯冬令营的日程表,预备再整装出发:去上课吟诗,去练习武术,去舞龙,去跳藏族的青春舞蹈,去品尝湖南佳肴,去观赏冬天奇景,去打雪仗……希望在中华文化弥漫的长沙天空再作一次深深的呼吸!看,我们都已陶醉在冬令营的激情里:望山,山还在招手;望湖,湖还在微笑;望你,你说这就是梦中宏伟的江山!望我,我口中的“乡愁”不再是“一湾浅浅的海峡”,而是“千里迢迢的故乡”。(长沙《湖南师范大学》特刊)
1.3.2 测定方法
普通光学显微镜下细胞计数板进行细胞计数,记数前在1 mL盐藻培养液中加入一滴鲁格氏液进行细胞固定。
细胞光密度于波长430 nm处测定,空白对照选用改良的Johnsons培养基。
色素测定:4 000 r/min离心5 min收集藻细胞,体积分数为80%丙酮重悬细胞后避光,4℃过夜提取色素后于波长470 nm、645 nm、662 nm处测定吸光度值,然后计算总胡萝卜素、叶绿素a与b的含量,计算公式如下[9-10]:

金属离子的测定采用原子吸收光谱法:4 000 r/min离心去除细胞,0.22 μm滤膜过滤,取7 mL去掉细胞的发酵液利用10 mL硝酸+2 mL高氯酸加热消化至溶液由黄变清体积大约2 mL后进行定容。金属离子钙(Ca)、钾(K)、镁(Mg)及铁(Fe)测定条件见表1。
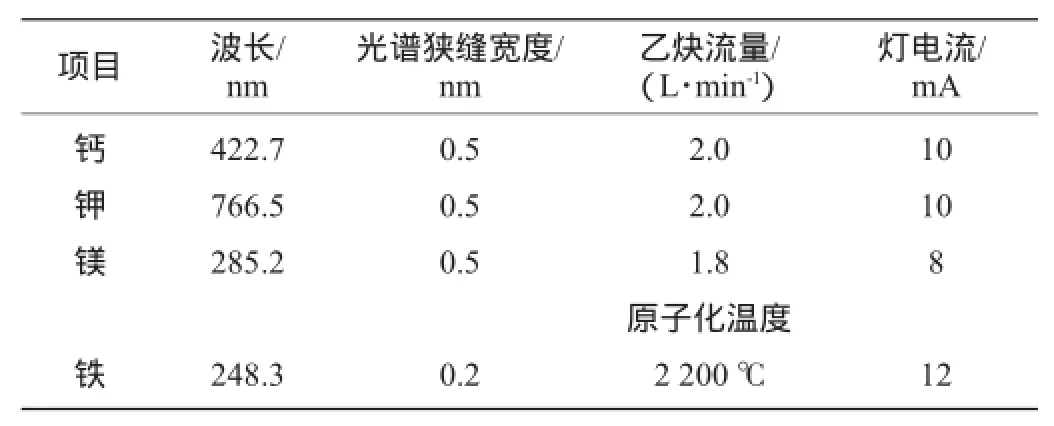
表1 原子吸收光谱采用的实验条件Table 1 Experimental conditions of atomic absorption spectrum
1.3.3 数据处理
采用Origin 8.5进行数据分析与作图;采用SPSS 19.0进行差异显著性分析。
2 结果与分析
2.1 杜氏盐藻钙、镁、钾、铁离子的消耗速率
在本研究的实验条件与接种密度下,杜氏盐藻的生长曲线见图1。由图1可知,杜氏盐藻生长的延滞期为3 d,在第4天进入对数生长期,在16 d后进入稳定生长期。
选择对数期的3个时间点(细胞密度为0.77× 106CFU/mL,1.84×106CFU/mL,2.12×106CFU/mL)与细胞密度不再继续增加的稳定期的两个时间点(4.52×106CFU/mL),测定以上5个时间点培养基中钙、镁、钾、铁离子的浓度。实验结果表明,随着细胞密度的升高,钾、镁离子在不同细胞密度时差异不显著(P>0.05),因此表明这两种离子的消耗速率近似为零。钙、铁离子在不同细胞密度的变化,结果见图2。由图2可知,钙、铁离子的消耗与细胞密度的增加呈现良好的线性关系,铁离子与钙离子在对数期前三个时间点的浓度值的线性拟合曲线的相关系数达0.999与0.895,拟合曲线分别为y=-42.46x+249.1与y=-0.708x+9.602。以增殖106个细胞消耗的离子量为计算单位,铁离子在对数期中的消耗速率为0.042 μg/106个细胞,钙离子在对数期中的消耗速率为0.708 μg/106个细胞。

图2 发酵液中铁离子(A)及钙离子(B)浓度的变化Fig.2 Changes of iron ion(A)and calcium ions(B)concentration at different growth phases
2.2 铁、钙离子浓度限制型培养基补料培养
在接种光密度(OD430nm)为0.1时,铁、钙离子浓度限制型培养基中含有的铁、钙离子的量够细胞长到细胞密度大约为3×106CFU/mL(OD430nm=2.0)。结果表明,最终细胞密度为3.88×106CFU/mL(OD430nm=2.47)。细胞进入稳定期后,继续补加细胞增加0.5×106CFU/mL细胞密度的铁与钙离子的量,结果表明细胞密度继续增加至4.22×106CFU/mL后停止增加(OD430nm=2.8),对比铁、钙离子前后对数期的杜氏盐藻细胞的光合速率、呼吸速率、光合色素含量,结果见表2。
由表2可知,铁、钙离子限制型培养基中藻细胞的净光合速率、叶绿素a、叶绿素b均显著低于完全培养基对照,两种培养基中的藻细胞的呼吸速率没有显著差异。铁、钙离子限制型培养基中藻细胞的总类胡萝卜素含量要高于完全培养基对照。

表2 离子限制型培养基铁、钙补料前后杜氏盐藻光合速率与色素含量Table 2 Comparisons of photosynthetic rates,pigments contents before and after ion feeding in ion type restrictions medium
2.3 培养基回流培养中铁、钙离子补料培养
铁、钙回流培养基中含有可供盐藻细胞长到密度4.62×106CFU/mL(OD430nm=3.0)的铁、钙离子的量。取对数期细胞接种于铁、钙回流培养基中,可得藻细胞的生长曲线见图3,与改良的Johnsons培养基中藻细胞相比,回流培养基中藻细胞没有延滞期,而改良的Johnsons培养基中藻细胞第3天细胞密度开始升高。由图3可知,回流培养基中的细胞在16 d到达稳定期,而改良的Johnsons培养基在20 d到达稳定期,细胞密度也大于回流培养基中的细胞。

图3 回流培养基与Johnsons培养基中杜氏盐藻的生长曲线Fig.3 Growth curves ofDunaliella salinain fed-batch media and Johnsons media
对比两种培养基中藻细胞对数期的光合作用于与色素组成,结果见表3。由表3可知,回流培养基藻细胞的净光合速率低于改良的Johnsons培养基中的,而呼吸速率没有显著地差异(P>0.05),回流培养基藻细胞的叶绿素a与b的含量均小于改良的Johnsons培养基中的含量,但总类胡萝卜素含量高于改良的Johnsons培养基中的含量。

表3 回流培养基与Johnsons培养基中杜氏盐藻光合速率与色素含量Table 3 Photosynthetic rates,pigments contents ofDunaliella salina in fed-batch media and Johnsons media
3 结论
研究微藻培养中矿质营养盐的消耗速率对解析微藻细胞的矿质营养机理与生产上的补料培养具有重要价值。杜氏盐藻是少数几种已经大规模培养的经济藻种之一,产生的胡萝卜素主要用于食品、化妆品以及医药领域,现在全球年产大约1 200 t,仍存在巨大的市场空缺[11]。已有研究表明,营养缺乏对于杜氏盐藻胡萝卜素的积累具有关键作用[7,12],因此研究各矿质元素的消耗速率对于实现通过控制营养盐的量偶联光温条件控制胡萝卜素积累的时间与含量的目标具有重要的意义。研究盐藻细胞对矿质营养元素的消耗速率相比于直接研究盐藻细胞的矿质元素组成具有明显的优势,如可以避免细胞清洗过程中对表面吸附离子的丢失而导致的实验误差等[13]。本研究采用每增殖106个细胞消耗的离子量来表征杜氏盐藻对金属离子的消耗速率相比用单位时间消耗的离子量来表征消耗速率,可以规避不同培养条件下生长速率的不同而产生的数据使用上的限制。钾镁离子为培养液中的大量元素,但杜氏盐藻对于这两种离子的利用量极低,超出了仪器的检测限制。本研究结果表明,在细胞进入稳定期后,杜氏盐藻细胞仍在继续吸收铁、钙离子,可能的原因是杜氏盐藻细胞为生长条件的好转做积累矿质营养。在对硝酸根与磷酸根阴离子的研究中(未发表数据),在杜氏盐藻细胞到达稳定期后,培养液中硝酸根与磷酸根的含量不再降低,说明杜氏盐藻细胞对铁、钙离子的利用上与对阴离子的利用方式存在不同。
铁、钙离子浓度限制型培养基中含有的铁、钙离子的量够细胞长到细胞密度OD430nm2.0,但结果表明,盐藻细胞密度最终达到OD430nm2.47,可能的原因是细胞铁、钙离子的储存作用,与盐藻细胞到达稳定期后铁、钙离子的继续降低数据相一致[14-15]。盐藻细胞的光合活性与色素含量相应减小说明铁、钙离子的浓度高低对藻细胞的光合作用系统有影响。回流培养基中藻细胞的延滞期消失说明回流培养基中的一些成分可以促使藻细胞对新培养环境的适应,但最终细胞密度没有达到理论上的OD430nm2.0,说明回流培养液经过多轮的利用后可能含有一些抑制细胞密度升高的成分。
[1]PRIETO A J,CANAVATE P,GARCIA-GONZALEZ M.Assessment of carotenoid production byDunaliella salinain different culture systems and operation regimes[J].J Biotechnol,2011,151(2):180-185.
[2]GOMEZ P I,GONZALEZ M A.The effect of temperature and irradiance on the growth and carotenogenic capacity of seven strains ofDunaliella salina(Chlorophyta)cultivated under laboratory conditions[J].Biol Res, 2005,38(2-3):151-162.
[3]BEN-AMOTZ A,AVRON M.On the factors which determine massive β-carotene accumulation in the halotolerant algaDunaliella bardawil[J]. Plant Physiol,1983,72(3):593-597.
[4]SALGUERO A,DE LA MORENA B,VIGARA J,et al.Carotenoids as protective response against oxidative damage inDunaliella bardawil[J]. Biomol Eng,2003,20(4):249-253.
[5]TSAI C F,LU F J,HSU Y W.Protective effects ofDunaliella salina-a carotenoids-rich alga-against ultraviolet B-induced corneal oxidative damage in mice[J].Mol Vis,2012,18(13):1540-1547.
[6]ZHANG S,LI X R,XU H,et al.Molecular cloning and functional characterization of MnSOD fromDunaliella salina[J].J Basic Microbiol, 2013,54(5):438-447.
[7]COESEL S N,BAUMGARTNER A C,TELES L M,et al.Nutrient limitation is the main regulatory factor for carotenoid accumulation and for Psy and Pds steady state transcript levels inDunaliella salina(Chlorophyta)exposed to high light and salt stress[J].Mar Biotechnol(NY),2008, 10(5):602-611.
[8]汪本凡.杜氏盐藻纯化及生物学特性研究[D].合肥:安徽农业大学硕士论文,2004.
[9]PISAL D S,LELE S S.Carotenoid production from microalga,Dunaliella salina[J].Indian J Biotechnol,2005,4(10):476-483.
[10]LICHTENTHALER H K,WELLBURN A R.Determination of total carotenoids and chlorophylls A and B of leaf in different solvents[J]. Biol Soc Trans,1985,11:591-592.
[11]U.S.Department of energy,office of energy efficiency and renewable energy.Biomass program[R].U.S.National Algal Biofuels Technology Roadmap(R),2010:62-63.
[12]SAHA S K,MOANE S,MURRAY P.Effect of macro-and micro-nutrient limitation on superoxide dismutase activities and carotenoid levels inmicroalgaDunaliellasalinaCCAP19/18[J].Bioresour Technol,2013, 147:23-28.
[13]唐颖,王长海,黄笛.无菌与带菌盐藻矿质元素含量的比较分析[J].光谱学与光谱分析,2010,30(7):1956-1959.
[14]记明侯.海藻化学[M].北京:科学出版社,1997.
[15]张宪政,陈凤玉,王荣富.植物生理学实验技术[M].辽宁:科学技术出版社,1994.
Effects of the fed-batch cultivation of mineral nutrients onDunaliella salinagrowth
LÜ Hexin,JIA Shiru*,CUI Xianggan,XIA Feng
(Tianjin Key Laboratory of Industrial Microbiology,Tianjin University of Science&Technology,Tianjin 300457,China)
The consumption rate ofDunaliella salinaon iron,potassium,calcium and magnesium were determined by atomic absorption spectrum. The metal ion content limit type medium was established and the effect of adding metal ion onD.salinagrowth,photosynthetic rate and pigment content were studied.The results showed that the consumption of iron and calcium decreased linearly during the log growth phase.The consumption rates of iron and calcium in the medium were 0.042 μg and 0.708 μg for 106cell proliferation,and after stable phase,calcium,iron concentration continued to fall.However,the difference of potassium and magnesium concentration in the medium was not significant in the growth period.The feeding of iron and calcium ions could promote the growth ofD.salinacells to revive but with a lower photosynthetic rates and chlorophyll contents, the total carotenoid content was higher than logarithmic phase before feeding.After recycling adding limited iron and calcium to the medium in the stable phase,the cell lag phase disappeared but the cell concentration decreased in the stable phase.The photosynthetic rate and chlorophyll content of cell in log phase in backflow medium decreased,but the total carotenoid content increased inversely.
Dunaliella salina;β-carotene;metal ion;consumption rate;consumption kinetics
Q939.1
A
0254-5071(2014)11-0051-04
10.11882/j.issn.0254-5071.2014.11.011
2014-09-17
中国博士后科学基金(2014M561189);长江学者和创新团队发展计划资助(IRT1166)
吕和鑫(1980-),男,助理研究员,博士,研究方向为微藻生物工程。
*通讯作者:贾士儒(1954-),男,教授,博士,研究方向为生物反应工程。
