两株H6N8亚型禽流感病毒分离株的分离与鉴定
2013-08-30崔鹏飞朱占松李文辉关立峥邓国华陈化兰
张 曦,崔鹏飞,朱占松,谭 丹,李文辉,关立峥,邓国华,陈化兰
(中国农业科学院哈尔滨兽医研究所农业部动物流感重点开放实验室/兽医生物技术国家重点实验室,黑龙江哈尔滨 150001)
野生水禽被认为是A型禽流感病毒(Avain influenza virues,AIV)的天然宿主和天然储存库,感染AIV的野生水禽在迁徙过程中通过排毒污染水源,从而感染其他动物,形成现有的流感病毒的传播生态[1]。自然情况下A型流感的宿主范围有一定特异性,新宿主出现或原宿主组成比例的变化都会改变病毒的生态并影响病毒的进化。作为宿主,家鸭在野生水禽向家禽传播流感的过程中起到关键作用[2-4]。中国在过去的20年里,家鸭养殖数量持续增加,目前国内家鸭数量占了全世界75%[5]。2000年~2005年间,3个主要的H6亚型AIV分支在国内建立,并在家鸭群体中传播[6-8]。为研究H6亚型AIV在家鸭群体中的传播,以及病毒在宿主体内的重组变异。本研究对我国南方活禽市场中的分离的两株鸭源H6N8亚型AIV进行全基因序列分析和动物致病性实验,并研究其相关的生物学特征。
1 材料和方法
1.1 病毒株和主要试剂 两株H6N8的AIV分离株均来自2011年华南地区活禽市场的正常家禽监测样品,由国家禽流感参考实验室分离保存,病毒命名为:A/duck/Fujian/S2191/2011(H6N8)(FJ/191/11);A/duck/Guizhou/S4035/2011(H6N8)(GZ/035/11)。10日龄SPF鸡胚及4周龄SPF鸡均购自哈尔滨兽医研究所实验动物中心;6周龄雌性BALB/c小鼠购自北京维通利华实验动物有限公司;RNA提取试剂TRIzol LS购自Invitrogen公司;DNA聚合酶和Marker 2000均购自TaKaRa公司;胶回收试剂盒购自上海华舜生物技术有限公司;反转录酶(MLV)试剂盒购自Invitrogen公司。
1.2 病毒的增殖及鸡胚半数感染量(EID50)测定 分离株经过有限稀释后接种10日龄SPF鸡胚纯化3代,进行病毒扩增;增殖后的H6N8亚型AIV按10倍倍比稀释,各稀释度经尿囊腔接种4枚10日龄SPF鸡胚,37℃培养,48 h后收集尿囊液测其血凝价,根据Reed-Muench法计算EID50。
1.3 基因组分析
1.3.1 RNA提取与PCR扩增采用TRIzol试剂盒,按说明书的步骤进行病毒RNA的提取,反转录后用于PCR,扩增产物按照胶回收试剂盒进行纯化。
1.3.2 序列测定与分析用BigDye Term inator测序试剂盒3.1version,按照测序试剂盒说明书进行测序。利用DNAStar软件中Seqman拼接序列,应用MegA lign(By Clustal V Method)进行同源性比较和进化分析,MEGA5(Construct/Test maximum likelihood Tree)绘制各基因节段的进化树。
1.4 抗原分析
1.4.1 制备免疫血清所需病毒SPF鸡胚接种48 h,收取鸡胚尿囊液,浓缩至血凝价7 log2~8 log2后与弗氏不完全佐剂1∶1混合,乳化后于鸡肌肉多点注射免疫。一个月后采血分离血清。
1.4.2 交叉血凝抑制(HI)试验利用病毒及其特异性血清,按照OIE颁布的标准HI方法,进行交叉HI试验。
1.4.3 抗原差异性分析抗原差异r值的计算方法按照文献[9]的方法进行。
1.5 动物感染试验
1.5.1 SPF鸡感染试验按每只106EID50/100μL的病毒鼻腔接种13只SPF鸡,另两只注射PBS作为对照。感染后连续8 d采集泄殖腔和喉头拭子,并接种鸡胚进行病毒分离、病毒滴度测定,分析其排毒规律。感染3 d后每株病毒采取3只鸡的组织脏器(脑、胸腺、气管、心、肝、脾、肺、肾、盲肠扁桃体、法氏囊)用于病毒分离滴定,病毒感染21 d后采集血清,检测血清阳转情况。
1.5.2 BALB/c小鼠感染试验按每只106EID50/50μL的病毒鼻腔接种8只小鼠,接种后第3 d迫杀3只,采集脑、鼻甲、脾、肾和肺用于病毒分离滴定;其余5只连续观察14 d,并记录体重变化情况。
2 结 果
2.1 序列及进化分析
2.1.1 HA基因的序列及进化分析AIV FJ/191/11和GZ/035/11分离株HA基因全长1 701 nt,编码566个氨基酸,HA裂解位点序列均为339PQIETR↓GLFG348,具有低致病性AIV的特征。两株病毒HA基因同源性差异较大,同源率为85.5%。两株病毒在第26位、27位、39位、306位和311位存在潜在糖基化位点,182位不存在潜在糖基化位点(表1)。FJ/191/11株HA基因属于ST2853-like分支,GZ/035/11株属于HuN573-like分支,均属于欧亚分支(图1)。
2.1.2 NA基因的序列及进化分析FJ/191/11和GZ/035/11分离株NA基因全长1 413 nt,NA基因同源率为76.4%,亲缘关系较远。与FJ/191/11株NA节段最接近的是A/duck/Thailand/CU-7518C/2010(H3N8)(JX307136.1),同源率为99%。与GZ/035/11株NA片段同源性最高的是A/swan/Shimane/42/1999(H7N8)(AB303079.1),同源率为94%。同源性高的节段并不局限于H6亚型AIV,同时还有H3和H7亚型AIV的N8节段(图1)。
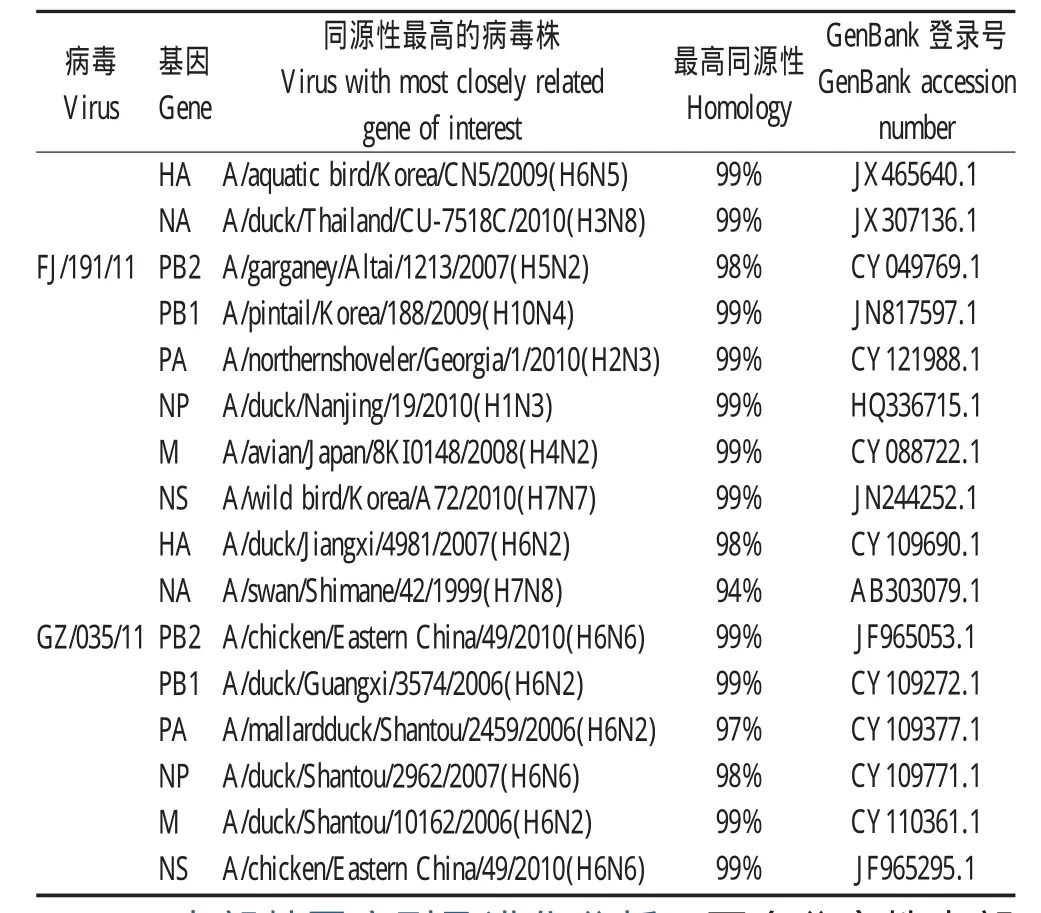
表1 毒株FJ/191/11及GZ/035/11的各基因片段BLAST分析结果Table 1 The blast analysis of genes of FJ/191/11 and GZ/035/11 influenza virus isolates
2.1.3 内部基因序列及进化分析两个分离株内部基因PB2、PB1、PA、NP、M、NS的核苷酸同源性分别为89.1%、90.0%、90.7%、94.3%、93.2%、93.3%。GZ/035/11中的PB2、PB1、PA、M、NS内部基因属于ST339-like分支,而FJ/191/11内部基因均属于HuN573-like分支,两株病毒NP基因则都属于HuN573-like分支。表明这两株病毒除了N8NA和NP基因以外,均有来自与不同的H6亚型分支,为H6家系内的病毒相互重组产生(图1)。
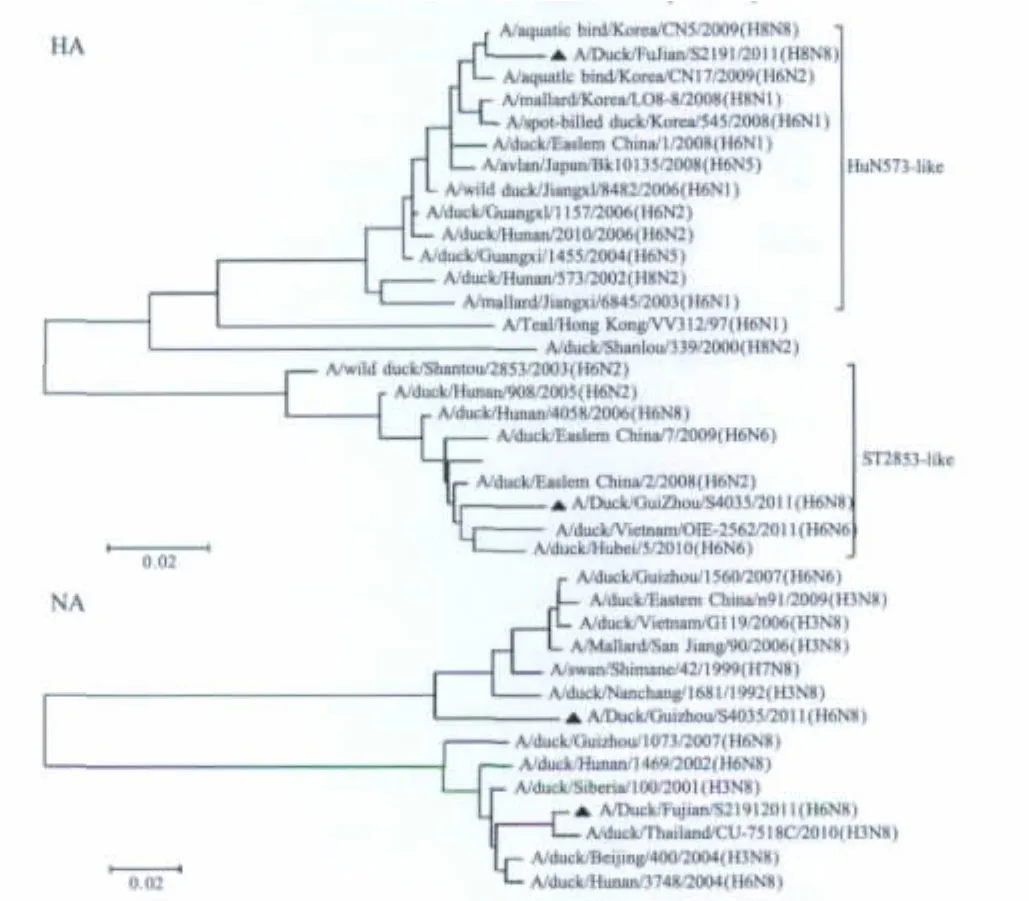
图1 FJ/191/11株和GZ/035/11株的基因进化树Fig.1 Phyogenetic tree based on HA and NA genes of FJ/191/11 and GZ/035/11
2.2 病毒对SPF鸡的感染性试验 两株病毒感染SPF鸡后均无明显临床症状,排毒方式有明显差异。感染21 d后血清检测结果显示:接种FJ/191/11病毒株的鸡血清转阳比例为8/9,接种GZ/035/11血清转阳比例为9/9,而且同居对照组鸡血清均呈阴性反应。病毒滴定结果显示:接种FJ/191/11病毒株的鸡群在泄殖腔排毒持续到第7 d,高峰期在第5 d,喉拭子病毒滴定结果为阴性;而接种GZ/035/11病毒株的鸡群,喉拭子排毒期只持续到第5 d,而泄殖腔拭子病毒滴定结果为阴性。同居鸡的拭子滴定结果和血清检测结果一致,表明此两株病毒均不能在鸡群之间发生有效传播(表2)。肺脏、气管、法氏囊、肾脏、盲肠扁桃体的脏器病毒检测结果均为阴性,表明两株病毒在所有脏器内均不能有效复制。
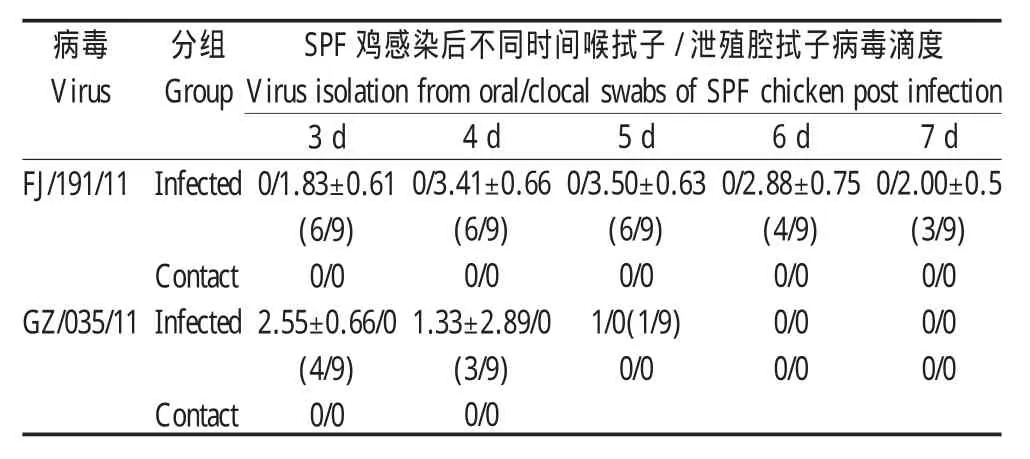
表2 病毒感染后的滴定结果Table 2 Virus titration after infection
2.3 病毒对BALB/c小鼠感染试验 106EID50鼻腔接种后,感染小鼠体重呈一过性下降,无明显临床变化。脏器病毒滴定结果显示,接种两株病毒的小鼠肺脏和鼻甲均能够分离到病毒,而其他脏器病毒滴定均为阴性(表3)。
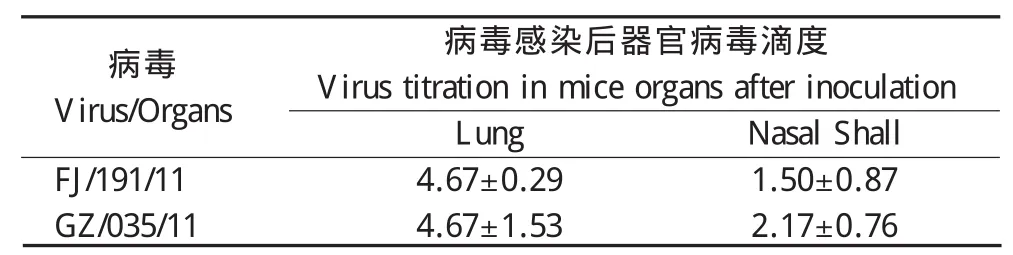
表3 感染后第3 d小鼠脏器病毒滴定Table 3 Virus titration of 2 H6N8 virues inmice organs on day 3 post inoculation
2.4 脏器的组织化学观察 病毒感染后3 d,两个感染组分别迫杀3只小鼠,采肺脏做组织病理变化检查,结果显示:小鼠肺脏局部呈现急性肺炎症状,表现为局部肺组织水肿,肺泡腔内出现炎性细胞的侵润,同时肺泡腔内的炎性细胞内出现阳性的抗原反应(图3)。
2.5 抗原分析结果 以两个分离株各自灭活病毒免疫鸡制备的特异性血清进行交叉HI试验,结果显示:FJ/191/11病毒株分别与两株病毒的特异性血清的HI血凝抑制价相差4 log2,GZ/035/11病毒株分别与两株病毒的特异性血清的HI血凝抑制价相差3 log2。两株病毒的抗原性相差系数r值为0.577,表明两株病毒抗原性差异较小。
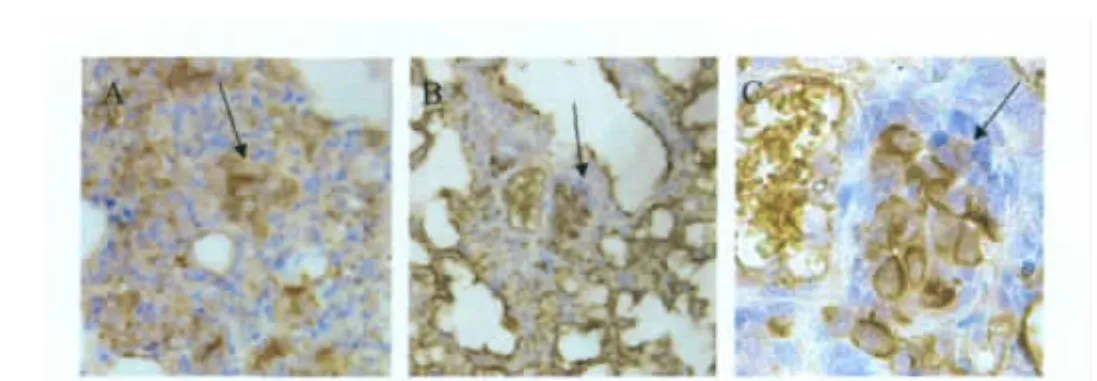
图3 感染FJ/191/11和GZ/035/11的免疫组织化学观察Fig.3 Immunohistochem ical observation of lung tissues in Mice infected FJ/191/11 and GZ/035/11
3 讨 论
本研究对2011年在我国华南地区活禽市场分离到的两株鸭源H6N8亚型AIV株的遗传演化分析表明,除N8NA基因以外的基因节段分别属于国内流行的H6家系,其中GZ/035/11病毒株的基因均属于ST339-like分支,而FJ/191/11病毒株基因均属于HuN573-like分支。在SPF鸡的感染性试验中,多脏器病毒滴定均为阴性,表明病毒不能在体内有效复制,并且不能发生水平传播感染;而两株病毒感染SPF鸡后排毒规律呈现明显的差异,FJ/191/11株感染组在泄殖腔中排毒,并能达到较高的病毒滴度;GZ/035/11株感染组则不能在泄殖腔中检测到病毒,表明两株病毒在SPF鸡体内对脏器的具有特异性的亲嗜性,不同来源的基因组成对病毒在禽类宿主中脏器的分布起到一定影响[10]。在小鼠的感染性试验中,尽管两株病毒的感染组小鼠的肺脏病毒滴定均为阳性,然而免疫组化显示,肺泡腔中的炎性细胞中抗原反应为阳性,这与可观察到的急性肺炎症状相符,也表明小鼠肺脏中的病毒在接种后3d并未清除完全。
尽管有研究显示H6亚型AIV能够与其他亚型内部基因HPAIV发生重组,本实验分离得到的病毒内部基因源于H6家系,不包含其他亚型AIV提供的内部基因;实验也证明两株H6N8病毒并没有在小鼠体内增殖的能力;但在SPF鸡感染试验中,能够不同程度排毒,表明H6N8亚型LPAIV有能力在水禽和家禽间跨种传播,增加鸭源H6亚型AIV在种家禽之间发生重组和传播的机会。同时,家鸭作为水禽与候鸟接触密切,在禽流感传播过程中起重要的作用[11],开展对鸭群的禽流感监测,对及时了解家禽AIV的感染情况,深入研究AIV传播的复杂性和多样性,都具有十分重要的意义。
[1]Webster R G,Bean W J,Gorman O T,et al.Evolution and ecology of influenza A viruses[J].M icrobiol Rev,1992,56:152-179.
[2]Laver W G,Webster R G.Studies on the origin of pandemic influenza.3.Evidence implicating duck and equine influenza viruses as possible progenitors of the Hong Kong strain of human influenza[J].Virology,1973,51(2):383-391.
[3]Sharp G B,Kawaoka Y,Webster R G.Coinfection of w ild ducks by influenza A viruses:distribution patterns and biological significance[J].JVirol,1997,71(8):6128-6135.
[4]Van Boeckel T P.Modelling the distribution of domestic ducks in Monsoon Asia[J].Agric Ecosyst Environ,2011,141:373-380.
[5]Cheung C L,Chen H,Guan Y,et al.Establishment of influenza A virus(H6N1)in Minor poultry species in southern China[J].JVirol,2007,81(19):10402-10412.
[6]Huang K,Peiris JS,Guan Y,et al.Establishment of an H6N2 influenza virus lineage in domestic ducks in southern China[J].JVirol,2010,84(14):6978-6986.
[7]Huang K,Peiris M,Guan Y,et al.Establishment and lineage replacement of H6 influenza viruses in domestic ducks in southern China[J].JVirol,2012,86(11):6075-6083.
[8]罗维玉,胡永浩,邓国华,等.两株鹅源H6N2亚型禽流感广东分离株的全序列分析及致病性研究[J].中国预防兽医学报,2012,34(5):345-349.
[9]Vanderven H A,Webster R G,Magor K E,et al.Avian influenza rapidly induces antiviral genes in duck lung and intestine[J].Mol Immunol,2012,51:316-324.
[10]Hulse-Post D J,Peiris JS,Webster R G,et al.Role of domestic ducks in the propagation and biological evolution of highly pathogenic H5N1 influenza viruses in Asia[J].Proc Natl Acad Sci,2005,202(30):10682-10687.
[11]Kim JK,Forrest H L,Webster R G,et al.Ducks:the"Trojan horses"of H5N1 influenza[J].Influenza Other Respi Viruses,2009,3(4):121-128.
