嗜盐古菌噬菌体研究进展
2012-01-12梅运军沈萍陈向东
梅运军,沈萍,陈向东*
(1.武汉工业学院化学与环境工程学院,湖北武汉430023;2.武汉大学生命科学学院病毒学国家重点实验室,湖北武汉430072)
噬菌体是感染细菌、真菌、放线菌或螺旋体等微生物的病毒的总称。据统计,截止2009年,已对5 500多株噬菌体进行了电镜观察,隶属于1目,17科[1]。近年的研究表明,噬菌体是一个庞大的生物群体,其总量超过了1030,并广泛分布于地球的各种环境中;而对古菌噬菌体的研究至今知之甚少,目前仅有50多株古菌噬菌体被报道,这些古菌噬菌体绝大多数来源于嗜热或极端嗜盐的环境中[2]。对于嗜盐古菌噬菌体的研究起源于上世纪70年代,Hs1是第1株被报道的嗜盐古菌噬菌体,随后Zillig及其同事对嗜盐古菌噬菌体ΦH进行了深入的研究,Dyall-Smith及Bamford领导的课题组在嗜盐古菌噬菌体研究方面也作出了巨大的贡献。然而,与细菌噬菌体研究相比较,对嗜盐古菌噬菌体的研究尚处在起步阶段,表1列出了部分研究较为深入的嗜盐古菌噬菌体。本文简要综述了嗜盐古菌噬菌体的形态及特性、分子生物学研究、嗜盐古菌噬菌体的分离及多样性研究方法等方面的进展。
1 嗜盐古菌噬菌体形态及特性
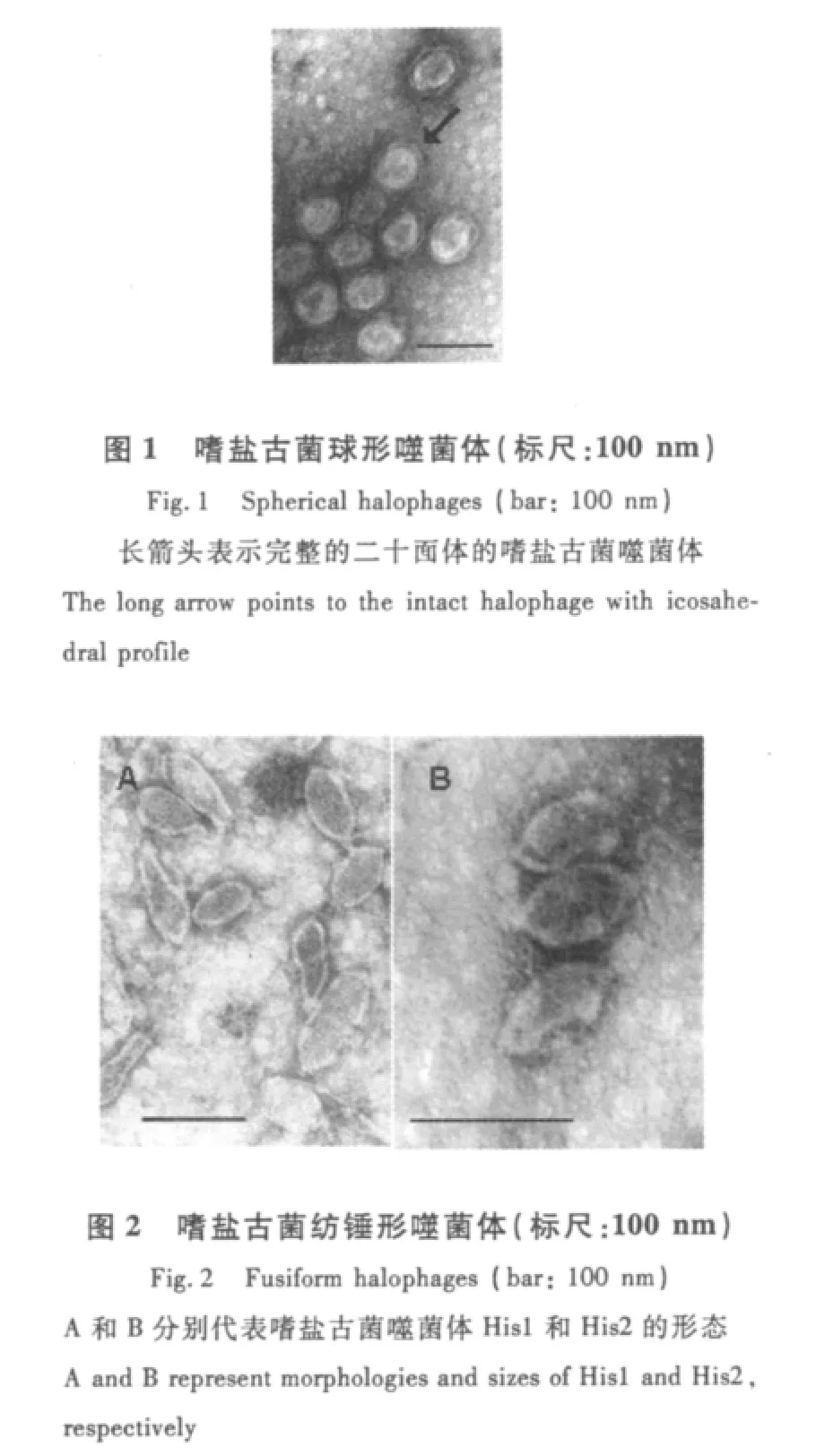
1.1 球形噬菌体
SH1为1株典型的嗜盐古菌球形噬菌体,如图1所示。其直径大约为70 nm,能感染宿主Haloarcula hispanica并形成清晰的噬菌斑[3],密度为1.33 g/mL,蛋白质衣壳由15种蛋白组成,分子量4~185 ku。就其外形而言,SH1类似于其他古菌噬菌体(PSV、TTSV1、STIV)以及细菌噬菌体PRD1,但几者之间存在一定的区别。其中,PSV和TTSV1具有相似的形态和基因组特征,核衣壳被脂膜外壳包裹,为双链线性DNA,PSV基因组末端具有反向重复序列,2条DNA链在末端共价结合[4]。SH1与STIV均为二十面体球状线性双链DNA噬菌体,与PSV、TTSV1不同的是SH1、STIV内部有一脂质层[5]。SH1与PRD1最为相似,不仅表现在蛋白质衣壳内有一脂质层,基因组为线性双链DNA,而且基因组大小和组织结构也极为相似[6]。

表1 部分嗜盐古菌噬菌体特性Table 1 Summary of partial halophages characteristics
1.2 纺锤形噬菌体
纺锤形(也称柠檬形)噬菌体是指一类形似纺锤、一端或两端有短尾的噬菌体。就目前已报道的纺锤形噬菌体而言,均来自泉古生菌界和广古生菌界。其中,His1和His2是能感染同一嗜盐古菌Haloarcula hispanic且一端具有短尾的纺锤形噬菌体,如图2所示,在分类上归于Salterprovirus属。His1(44 nm×77 nm)与His2(44 nm×67 nm)大小相似,密度相近(His1ρ=1.28 g/mL,His2ρ=1.30 g/mL),基因组皆为线性双链DNA,His1基因组由14 464 bp组成,His2由16 067 bp组成,G+C的摩尔百分比为40%,基因组末端具有反向重复序列并含有末端结合蛋白,缺少DNA整合酶编码基因[7]。尽管两者如此相似,但在基因组的碱基排列上存在很大的差别;在蛋白质(经开放阅读框预测)水平上除了DNA聚合酶具有相似的氨基酸残基外(42%的一致性),其他预测蛋白都不具有明显的相似之处。His2噬菌体颗粒衣壳蛋白由4种主要的蛋白组成—VP1、VP2、VP3、VP4,其大小分别为62、36、28、21 ku。
1.3 头尾形噬菌体
典型的头尾形噬菌体是由二十面体的头部以及螺旋对称的可伸缩或不可伸缩的尾部组成,尾部包括尾髓、尾鞘、尾板、尾刺和尾丝。此类噬菌体广泛存在于嗜盐古菌中,包括Hh1、Hh3、Ja.1、Hs1、ΦH、ΦN、ΦCh1、S45、S5100、B10、SNJ1、HF1、HF2等,其中,ΦH、ΦCh1、S5100、HF1、HF2具有明显可伸缩的尾部,Hh1、Hh3、Ja.1、Hs1、ΦN、S45、B10、SNJ1尾部不可伸缩,典型的嗜盐古菌头尾形噬菌体代表如图3所示。除基因组未知的嗜盐古菌噬菌体B10、S45、Ja1外,其余基因组皆为线性、双链DNA,有趣的是在ΦCh1蛋白质衣壳内还包含有一段宿主的RNA核苷酸序列,大小在80~700 bp。在分类学上,ΦCh1、HF1、HF2属于Myoviridae科。

图3 嗜盐古菌头尾形噬菌体(标尺:100 nm)Fig.3 Head-tail halophages(bar:100 nm)
1.4 其他形状的噬菌体
除了常见的3种形状的噬菌体之外,在一些盐浓度较高的环境水样中,还存在其他形状的噬菌体颗粒,如发夹形、丝状、链状、钩状、蝌蚪状、芦苇状以及复杂结构的噬菌体颗粒[8],见图4。

图4 其他形状的噬菌体(标尺:100 nm)Fig.4 Other morphologies of halophages(bar:100 nm)
然而,对这些形态各异的噬菌体的研究目前仅局限于形态学的报道,其他方面的研究有待于进一步的深入。
2 嗜盐古菌噬菌体分子生物学研究
对嗜盐古菌噬菌体分子生物学的研究起源于上世纪80年代后期,ΦH为1株具有头尾结构的温和型噬菌体,基因组大小为59 kb双链DNA,除甲基化酶基因之外,该噬菌体基因组序列与细菌、噬菌体或真菌病毒的基因组序列很少有相似之处,直到另一株嗜盐古菌噬菌体ΦCh1基因组测序完成,两者在核苷酸序列上相似性为97%,而且在开放阅读框上噬菌体ΦCh1与ΦH也非常相近[9-10]。90年代初,Dyall-Smith及其研究团队在嗜盐古菌噬菌体方面做了大量的工作,发现并比较深入地研究了His1、His2、HF1、HF1和SH1,涉及噬菌体基因组组成及结构、衣壳蛋白及转录等各方面[3,6,11-16]。在基因结构及组成上以HF1、HF2为例,两者具有相似的基因组大小(大约77 kb),而且通过交叉杂交分析发现两者基因组序列具有80%一致性;基因组测序完成后,两者在序列上的异同更加清晰。

图5 HF1与HF2基因组的图形比对Fig.5 Graphical alignment of HF1 and HF2 genomes
图5示意了HF1与HF2基因组的结构以及两者基因组比对情况,图中两基因组示意图间的垂直短线表示两者基因组中存在的碱基替换的位置,深黑色和不连续线条分别表示插入和缺失的序列。显然,两者基因组前48 kb(大约基因组60%)除了1个碱基的差异外,其余部分完全一致,但是剩下部分的基因组有非常明显的差异,只有87%的一致性。研究认为,造成序列差异的原因主要是在嗜盐古菌中基因重组现象普遍存在,因此HF1与HF2序列间的差异可能是HF1或HF2与另外一类HF类似的噬菌体间发生了重组,导致部分序列被取代;而且HF1与HF2晚期表达的基因(如衣壳蛋白基因)存在于这些差异的序列中,致使衣壳蛋白存在差异,最终导致两者侵染宿主上的不同[17]。
在复制方式HF2类似于大肠埃希菌噬菌体T3、T7,复制过程中基因组形成串联中间体,在末端酶或包装酶作用下将病毒基因组从串联体上特异位点切割并进行核衣壳的包装;而SH1复制方式类似于腺病毒及噬肝DNA病毒,以蛋白质为引物引发DNA高效合成,但在整个噬菌体SH1的基因组中未发现编码DNA聚合酶的基因,因此推测该病毒的复制过程依赖宿主的DNA聚合酶[12]。
在转录方面,与细菌噬菌体相比,嗜盐古菌噬菌体也有许多特别之处,而古菌噬菌体间存在很多相似之处。对一般细菌噬菌体而言,转录分为前期转录、中期转录以及晚期转录3个阶段,而已报道的嗜盐古菌噬菌体SH1,通过RT-PCR分析转录分为早期和晚期过程,无明显的中期时相,这一现象在其他古菌噬菌体中也有类似的报道,如SIRV1和SIRV2[14,18];转录时,编码衣壳蛋白的转录本在噬菌体感染的前期就已经开始合成直至整个感染周期,只是这些结构蛋白基因以及包装ATP酶基因在感染的后期具有更高的转录[18]。在转录形式上,在噬菌体SH1的基因都以操纵子的形式被转录,类似于嗜热古菌噬菌体SSV1,而且存在转录终止子通读现象,在以往的RNA病毒以及古菌微卫星病毒pSSVx中都有类似的报道[19-21]。另外,在感染的后期非编码链有不同程度的转录,特别是出现转录通读后。这些非编码链转录形成的转录本并不编码功能性蛋白,推测在蛋白质翻译过程中起到翻译调控的作用,在早期的研究中,也发现了嗜盐古生菌噬菌体ΦH存在这种dsRNA和iRNA的调控作用,近年在古菌微卫星病毒pSSVx中也有类似的反义RNA的调控作用[21]。尽管对嗜盐古菌噬菌体分子生物学方面的研究取得了一定的成绩,但是与细菌噬菌体以及真核生物的病毒相比,尚存很大差异,还需要大量的基础研究。
3 嗜盐古菌噬菌体分离及噬菌体群体研究方法
嗜盐古菌噬菌体分离常采用如下3种方法:①实验室敏感菌株分离法;②基因组脉冲场凝胶电泳分离法;③相分配法。其中,实验室敏感菌株分离法是最常用的方法,就目前已报道的绝大部分嗜盐古菌噬菌体均采用这种方法;但该方法也存在一些缺陷,如仅对烈性噬菌体有效,对温和噬菌体却失去了优势,由于目前报道的可作为敏感菌株的宿主有限,因此该方法只能检测环境中的部分噬菌体[7,17]。基因组脉冲场凝胶电泳法是近来发展起来的一种分离环境样品中嗜盐古菌噬菌体基因组的方法,利用该方法虽然可以得到大量噬菌体的基因组序列,但噬菌体的形态以及噬菌体与宿主的动态关系却一无所知[22-23]。相分配法主要借助噬菌体颗粒自身物理及化学性质(如噬菌体颗粒大小或浮力密度等)而采取的必要的分离、纯化噬菌体的步骤,该方法也常用到分离其他噬菌体上,但不同的是在分离嗜盐古菌噬菌体时常采用高盐的缓冲液作为调节渗透平衡的体系。
噬菌体群体研究主要采用的手段包括:①过滤法对样品进行浓缩,结合透射电子显微镜或荧光电子显微镜对噬菌体形态进行观察[24-25];②脉冲场凝胶电泳法结合DNA杂交法对噬菌体基因组进行分析[23];③宏基因组法对环境样品进行噬菌体基因组多样性分析[26]。
4 问题与展望
对嗜盐古菌噬菌体的研究虽然取得了一定的成就,但与对细菌噬菌体研究相比尚处于研究的早期。在噬菌体形态多样性研究方面,就目前报道纺锤体形、球形及星形噬菌体十分普遍,而头尾噬菌体却很少,主要原因在于检测和分离方法不当,如主要采用在实验室利用有限的可培养的宿主菌对噬菌体分离(常利用Hbt.Salinarum作为宿主),忽视了环境中噬菌体的多样性;而对嗜盐古菌噬菌体分子生物学的研究也仅仅局限于极个别噬菌体,缺少噬菌体基因组学与蛋白质组学的大量研究,不利于对噬菌体起源以及生命起源与进化的分析;对于这类特殊的噬菌体在生态中的地位以及与宿主的相互关系的研究也十分的薄弱,缺乏较为系统的研究。总之,随着对嗜盐古菌噬菌体研究的深入,人们将逐渐加深对自然界中这类特殊的噬菌体的了解,并为自然界中生命的起源与进化提供依据。
[1] Ackermann HW.Phage classification and characterization[J].Methods in Molecular Biology,2009,501:127-140.
[2] Mochizuki T,Yoshida T,Tanaka R,et al.Diversity of viruses of the hyperthermophilic archaeal genus Aeropyrum,and isolation of Aeropyrum pernix baciliform virus 1,APBV1,the first representative of the familiy Clavaviridae[J].Virology,2010,402(2):347-354.
[3] Porter K,Kukkaro P,Bamford JKH,et al.SH1:A novel,spherical halovirus isolated from an Australian hypersaline lake[J].Virology,2005,335(1):22-33.
[4] Häring M,Peng X,Brügger K,et al.Morphology and genome organization of the virus PSV of the hyperthermophilic archaeal genera Pyrobaculum and Thermoproteus:a novel virus family,the Globuloviridae[J].Virology,2004,323(2):233-242.
[5] Maaty WSA,Ortmann AC,Dlaki M,et al.Characterization of the achaeal Thermophile Sulfolobus Turreted icosahedral virus validates an evolutionary link among double-strand DNA viruses from all domains of life[J].Journal of Virology,2006,80(15):7625-7635.
[6] Bamford DH,Ravantti JJ,Rönnholm G,et al.Constituents of SH1,a novel lipid-containing virus infecting the halophilic euryarchaeon Haloarcula hispanica[J].Journal of Virology,2005,79(14):9097-9107.
[7] Prangishvili D,Forterre P,Garrett RA.Viruses of the Archaea:a unifying view[J].Nature Reviews Microbiology,2006,4(11):837-848.
[8] Sime-Ngando T,Lucas S,Robin A,et al.Diversity of virushost systems in hypersaline Lake Retba,Senegal[J].Environmental Microbology,2011,13(8):1956-1972.
[9]Schnabel H,Zillig W,Pfaffle M,et al.Halobacterium halobium phage φH[J].EMBO J,1982,1(1):87-92.
[10] Klein R,Baranyi U,Rossler N,et al.Natrialba magadii virus φCh1:First complete nucleotide sequence and functional organization of a virus infecting a haloalkaliphilic archaeon[J].Molecular Microbiology,2002,45(3):851-863.
[11] Nuttall S,Dyall-Smith ML.HF1 and HF2:Novel bacteriophages of halophilic Archaea[J].Virology,1993,197(2):678-684.
[12] Nuttall S,Dyall-Smith ML.Halophage HF2:genome organisation and replication strategy[J].Journal of Virology,1995,69(4):2322-2327.
[13] Porter K,Dyall-Smith ML.Transfection of haloarchaea by the DNAs of spindle and round haloviruses and the use of transposon mutagenesis to identify non-essential regions[J].Molecular Microbiology,2008,70(5):1236-45.
[14] Porter K,Russ BE,Ji Y,et al.The transcription program of the protein-primed halovirus SH1[J].Microbiology,2008,154(11):3599-3608.
[15] Tang SL,Nuttall S,Dyall-Smith,M.Haloviruses HF1 and HF2:evidence for a recent and large recombination event[J].Journal of Bacteriology,2004,186(9):2810-2817.
[16] Tang S L,Nuttall S,Ngui K,et al.HF2:a double-stranded DNA tailed haloarchaeal virus with a mosaic genome[J].Molecular Microbiology,2002,44(1):283-296.
[17] Dyall-Smith M,Tang S,Batch C.Haloviruses;how diverse are they?[J].Research in Microbiology,2003,154(4):309-313.
[18] Kessler A,Brinkman AB,van der O,et al..Transcription of the rod-shaped viruses SIRV1 and SIRV2 of the hyperthermophilic archaeon Sulfolobus[J].Journal of Bacteriology,2004,186(22):7745-7753.
[19] Fröls S,Gordon PM,Panlilio M A,et al.Elucidating the transcription cycle of the UV-inducible hyperthermophilic archaeal virus SSV1 by DNA microarrays[J].Virology,2007,365(1):48-59.
[20] Reiter WD,Palm P,Yeats S,et al.Gene expression in archaebacteria:physical mapping of constitutive and UV-inducible transcripts from the Sulfolobus virus-like particle SSV1[J].Molecular and General Genetics,1987,209(2):270-275.
[21] Contursi P,Cannio R,Prato S,et al.Transcriptional analysis of the genetic element pSSVx:differential and temporal regulation of gene expression reveals correlation between transcription and replication[J].Journal of Bacteriology,2007,189(17):6339-6350.
[22] Santos F,Yarza P,Parro V,et al.Metagenomic approach to the study of haloviruses:the environmental halophage 1[J].Environmental Microbiology,2007,9(7):1711-1723.
[23] Santos F,Yarza P,Parro V,et al.The metavirome of a hypersaline environment[J].Environmental Microbiology,2010,12(11):2965-2976.
[24] Sime-Ngando T,Lucas S,Robin A,et al.Diversity of virushost systems in hypersaline Lake Retba,Senegal[J].Environmental Microbiololgy,2011,13(8):1956-1972.
[25] Brum JR,Steward GF.Morphological characterization of viruses in the stratified water column of alkaline,hypersaline Mono Lake[J].Microbial Ecology,2010,60(3):636-643.
[26] Angly LE,Willner D,Prieto-Davó A,et al.The GAAS metagenomic tool and its estimations of viral and microbial average genome size in four major biomes[J].PloS computational Biology,2009,5(12):e1000593.
