鸡传染性法氏囊病病毒超强毒株VP2基因突变对细胞嗜性改变的研究
2010-05-21祁小乐高玉龙高宏雷秦立廷邓小芸王笑梅
高 立,祁小乐,高玉龙,高宏雷,秦立廷,邓小芸,李 凯,王笑梅
(中国农业科学院哈尔滨兽医研究所兽医生物技术国家重点实验室/禽传染病研究室,黑龙江 哈尔滨 150001)
鸡传染性法氏囊病(Infectious bursal disease,IBD)是由IBD病毒(IBDV)引起的一种危害鸡的急性、高度接触性传染病[1]。超强致病性IBDV(vvIBDV)经传代致弱能够感染非允许细胞鸡胚成纤维细胞(CEF),通过病毒碱基突变不仅有助于了解IBDV的遗传变异机制,而且对于研发新型疫苗意义重大。近年来,IBDV遗传变异和细胞嗜性等分子基础研究取得了一些进展[2-4]。VP2被认为是决定病毒细胞嗜性的基因[5-6],然而,其具体的分子机制尚存在争议。我国在这方面的研究尚未见报道。本课题组利用已建立的IBDV反向遗传操作系统[7],对IBDV细胞嗜性的分子基础进行分析和预测。本研究借助该操作技术,对国内地方分离的vvIBDV,通过定点突变和重叠延伸PCR(SOE-PCR)等技术,研究定位vvIBDV的VP2基因突变对细胞嗜性改变的影响。
1 材料和方法
1.1 重组质粒及单克隆抗体(MAb) 重组质粒pT-HLJ0504A、pT-HLJ0504B(分别含有 vvIBDV分离毒株HLJ-0504全基因组的A、B节段)由本课题组构建保存;IBDV VP2 MAb由本课题组制备。
1.2 主要试剂 DpnⅠ酶购自NEB公司;Plasmid Midi Kit购自QIAGEN公司;LipofectamineTM2000和SuperscriptTMRT-PCR Systems试剂盒购自英俊生物技术有限公司;Opti-MEMⅠMedium购自GIBCO公司;FITC标记的羊抗鼠IgG购自Sigma公司。
1.3 引物设计与合成 实验中所用引物见表1,引物由英俊生物技术有限公司合成。
1.4 感染性分子克隆的构建
1.4.1 核酶结构的引入 参考文献[7]的方法,以AU01/AL01、AU02/AL02和AU02/AL03为引物,分3步在基因组A节段的5'端引入HamRz(5'-TGTT AAGCGTCTGATGAGTCCGTGAGGACGAAACTATA GGAAAGGAATTCCTATAGTC-3'),3'端引入 HdvRz(5'-GGGTCGGCATGGCATCTCCACCTCCTCGCGGT CCGACCTGGGCATCCGAAGGAGGACGCACGTCCACTCGGATGGCTAAGGGAGGGCG-3')。将最终PCR产物命名为HLJ0504AHRT。将其克隆于pMD18-T载体,重组质粒命名为pT-HLJ0504AHRT。用引物BU01/BL01、BU02/AL02和BU02/BL03以同样方法在基因组B节段两端引入核酶结构,将最终PCR产物命名为HLJ0504BHRT。
1.4.2 突变位点的引入 参考文献[7]中点突变方法,以pT-HLJ0504AHRT为模板,GxA889TU/Gx-A889TL,GxG980AU/GxG980AL为引物(表1)。在A节段上同时引入核苷酸突变A889T、G980A,即分别在VP2上引入氨基酸突变Q253H和A284T,突变后的重组质粒命名为pT-HLJ0504A889/980HRT。

表1 引入突变、核酶结构和RT-PCR的引物Table 1 Primers for mutation,ribozyme sequences introduction and RT-PCR
1.4.3 全基因组克隆的构建 用SacⅠ和KpnⅠ分别处理HLJ0504AHRT、pT-HLJ0504A889/980HRT和真核表达载体pCAGGS,纯化后分别连接。测序鉴定正确的重组质粒分别命名为pCAGGHLJ 0504AHRT、pCAGGHLJ0504A889/980HRT(图 1)。
用ClaⅠ和XhoⅠ处理HLJ0504BHRT,与经相同酶酶切处理的pCAGGS载体连接,将测序正确的重组质粒命名为pCAGGHLJ0504BHRT(图1)。
1.5 病毒拯救及鉴定 提取重组质粒,参考文献[7]的方法,将含A、B节段的重组质粒共转染DF1细胞进行病毒拯救,在CEF细胞上盲传直至产生细胞病变(CPE)。转染分两个组:pCAGGHLJ-0504AHRT和 pCAGGHLJ0504BHRT; pCAGGHLJ-0504A889/980HRT和pCAGGHLJ0504BHRT。同时设立只转染空载体pCAGGS作为空白对照。

图1 重组质粒示意图Fig.1 Schematic diagram of the recombinant plasmids
1.5.1 间接免疫荧光检测(IFA) 将拯救病毒感染CEF,16 h后无水乙醇固定细胞,1∶100稀释IBDVVP2 MAb为一抗,作用1 h。以1∶100稀释FITC标记的羊抗鼠IgG为二抗,作用1 h,PBST洗涤后荧光显微镜观察结果。同时设置阴性对照。
1.5.2 电镜观察 将收获的拯救病毒细胞悬液反复冻融3次,3000 r/min于4℃离心10 min,取上清12000 r/min于4℃离心10 min,适量PBS重悬沉淀,负染电镜观察。
1.5.3 RT-PCR鉴定 按SuperscriptTMRT-PCR Systems试剂盒说明书,取第4代细胞毒提取总RNA,以引物AU/F6VP2Q1512L和BU/B1344L分别RT-PCR扩增A、B节段的部分片段进行检测,并进行测序鉴定。
1.6 拯救病毒复制动力学研究 按常规方法滴定拯救病毒的TCID50效价。以104TCID50拯救毒接种CEF单层,分别于感染后每隔12 h取细胞上清,滴定其TCID50效价;重复测定3次,取其平均值,绘制病毒复制动力学曲线。
2 结 果
2.1 感染性分子克隆的构建 A、B节段经突变及引入核酶结构后,经PCR扩增分别获得约为3400 bp和3000 bp,与预期(3426 bp和2985 bp)结果相符(图 2)。构建的重组质粒 pCAGGHLJ-0504AHRT、pCAGGHLJ0504A889/980HRT经SacⅠ/KpnⅠ双酶切后获得片段,与预期大小相符(图略)。重组质粒pCAGGHLJ0504BHRT经ClaⅠ/XhoⅠ酶切鉴定完全正确(图略)。进一步的 pCAGGHLJ0504-AHRT、pCAGGHLJ0504A889/980HRT和pCAGGHLJ-0504BHRT克隆全长测序显示,双点突变、核酶结构引入及开放阅读框完全正确。
2.2 病毒拯救 pCAGGHLJ0504A889/980HRT与pCAGGHLJ0504BHRT共转染组,在CEF上传第2代时,可见极轻微CPE;第2代以后,均可见较明显CPE;接毒后72 h,细胞碎裂但不悬浮,折光性增强,呈砂砾状。pCAGGHLJ0504AHRT与pCAGGHLJ0504BHRT共转染组重复转染5次,并在细胞上连续盲传5代以上,均没有观察到任何CPE。将pCAGGHLJ0504A889/980HRT和 pCAGGHLJ0504-BHRT共转染获得的拯救毒命名为rHLJ-0504HT。
2.3 拯救毒的鉴定 rHLJ-0504HT感染CEF后IFA可检测到特异荧光信号(图3)。电镜下可观察到rHLJ-0504HT细胞培养物上清中有直径60 nm左右、无囊膜的病毒粒子,这与IBDV特有的形态结构一致(图略)。而 pCAGGHLJ0504AHRT和 pCAGGHLJ-0504BHRT共感染组的细胞悬液接种CEF后IFA为阴性,电镜下也没有观察到病毒粒子,没有获得拯救病毒。
2.4 拯救病毒RT-PCR鉴定 从rHLJ-0504HT第4代毒中扩增A、B节段部分片段,获得了预期大小约为1400 bp、1340 bp的片段(图略)。测序结果显示:A节段存在突变位点T889和A980,A、B节段的其余部分与亲本毒HLJ-0504序列完全一致。
2.5 复制动力学研究 依据感染CEF后不同时间段细胞培养液中病毒的TCID50效价,绘制了拯救病毒rHLJ-0504HT的复制动力学曲线(图4)。接毒后12 h可检测到病毒;24 h达到病毒复制平台期,平台期病毒的效价均在105TCID50/mL以上;在接毒的前48 h内,病毒的效价呈上升趋势,但是上升幅度较小;接毒60 h后,拯救病毒的效价开始有所下降,下降幅度也较小。


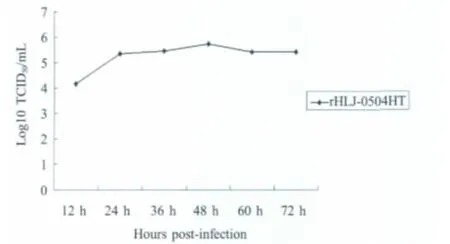
图4 拯救病毒rHLJ-0504HT复制动力学曲线Fig.4 The replication kinetics curve of the rescued rHLJ-0504HT
3 讨论
IBDV变异和细胞嗜性的分子基础研究表明,VP2决定IBDV的细胞嗜性[5-6]。通过定点突变,将影响IBDV细胞嗜性的分子基础进一步集中在了VP2高变区253位、284位、279位和330位氨基酸上。除330位氨基酸一致认为与IBDV细胞嗜性无关外,253位、284位和279位氨基酸与IBDV细胞嗜性的关系,则存在较大的异议[8-12]。为了进一步研究和定位vvIBDV细胞嗜性改变的分子基础,本研究利用反向遗传操作技术和定点突变技术,对vvIBDV HLJ-0504株进行了定点突变和病毒拯救。亲本病毒HLJ-0504由于不能适应CEF,共转染其基因组不能获得拯救病毒。而将其A节段VP2中的两个氨基酸位点进行突变(Q253H/A284T)后,与其B节段共转染可以获得拯救病毒。因此,双位点氨基酸突变改变了vvIBDV的细胞嗜性,使其适应CEF细胞。证明VP2的253位和284位氨基酸决定vvIBDV的细胞嗜性。
VP2的晶体结构显示,VP2亚单位分为3个明显的部分,即B(Bas)区、S(Shell)区和P(Projection)区,其中P突出于病毒粒子的最表层[13-14]。运用在线软件Swiss Model对IBDV的VP2蛋白进行同源建模分析发现:253和284两个氨基酸残基位于VP2蛋白的P(projection)区的突出部位,253位于PDE loop,284位于 PFG loop。PDE loop和 PFG loop被认为与病毒-受体结合密切相关[14]。其中VP2 Q253H的突变将使线性的氨基酸变为有环状结构的氨基酸,增加了β转角形成的可能性;而A284T的突变使非极性的疏水氨基酸变为极性的亲水氨基酸,增加了β折叠形成的可能性,而且还增加了一个酪蛋白激酶Ⅱ磷酸化潜在位点。所以253位和284位两个氨基酸残基的改变极有可能造成病毒配体分子结构的改变,进而改变病毒与宿主细胞受体的结合状态和类型,最终改变病毒的细胞嗜性。
本研究首次利用我国vvIBDV分离超强毒株,采用反向遗传操作系统证明:VP2的253位和284位氨基酸决定了IBDV的细胞嗜性。本研究不仅有助于了解IBDV的遗传变异机制,而且对于研发新型疫苗提供了实验依据。
[1]Mü ller H,Islam M R,Raue R.Research on infectious bursal disease-the past,the present and the future[J].Vet Microbiol,2003,97:153-165.
[2]王笑梅,王秀荣,陈冠春,等.鸡传染性法氏囊病超强毒致弱株的生物学特性研究[J].中国预防兽医学报,1999,21(6):436-438.
[3]Yu Wen Y Q,Gao Y L,Gao H L,et al.Sequence analysis of the VP2 hypervariable region of eight very virulent infectious bursal disease virus isolates from the Northeast of China[J].Avian Dis,2008,52:284-290.
[4]Brandt M,Yao K,Liu M,et al.Molecular determinants of virulence,cell tropism,and pathogenic phenotype of infectious bursal disease virus[J].J Virol,2001,75:11974-11982.
[5]Boot H J,Agnes H M,Arjan J W.Rescue of very virulent and mosaic infectious bursal disease virus from cloned cDNA:VP2 is not the sole determinant of the very virulent phenotype[J].J Virol,2000,74:6701-6711.
[6]Van Loon A A,De Haas N,Zeyda I,et al.Alteration of amino acids in VP2 of very virulent infectious bursal disease virus results in tissue culture adaptation and attenuation in chickens[J].J Gen Virol,2002,83:121-129.
[7]Qi X,Gao Y,Gao H,et al.An improved method for infectious bursal disease virus rescue using RNA polymerase II system[J].J Virol Meth,2007,142:81-88.
[8]Brandt M,Yao K,Liu M,et al.Molecular determinants of virulence,cell tropism,and pathogenic phenotype of infectious bursal disease virus[J].J Virol,2001,75:11974-11982.
[9]Lim B L,Cao Y C,Yu T,et al.Adaption of very virulent infectious bursal disease virus to chicken embryonic fibroblasts by site-directed mutagenesis of residues 279 and 284 of viral coat protein VP2[J].J Virol,1999,73:2854-2862.
[10]Mundt E.Tissue culture infectivity of different strains of infectious bursal disease virus is determination by distinct amino acids in VP2[J].J Gen Virol,1999,80:2067-2076.
[11]Van Loon A A,De Haas N,Zeyda I,et al.Alteration of amino acids in VP2 of very virulent infectious bursal disease virus results in tissue culture adaptation and attenuation in chickens[J].J Gen Virol,2002,83:121-129.
[12]Coulibaly F,Chevalier C,Gutsche I,et al.The birnavirus crystal structure reveals structural relationships among icosahedral viruses[J].Cell,2005,120:761-772.
[13]Garriga D,Querol-Audi J,Abaitua F,et al.The 2.6-Angstrom structure of infectious bursal disease virus-derived T1 particles reveals new stabilizing elements of the virus capsid[J].J Virol,2006,80:6895-6905.
[14]Lee C C,Ko T P,Chou C C,et al.Crystal structure of infectious bursal disease virus VP2 subviral particle at 2.6 A resolution:implications in virion assembly and immunogenicity[J].J Structural Biol,2006,155:74-86.
