微生物酪氨酸解氨酶的研究进展
2010-03-15张峰曾化伟廖祥儒蔡宇杰童超
张峰, 曾化伟, 廖祥儒, 蔡宇杰, 童超
(1.江南大学工业生物技术教育部重点实验室,江苏无锡214122;2.江南大学生物工程学院,江苏无锡214122)
次生代谢物质对医药、轻工、化工、农药等领域的发展具有不可替代的地位。其来源包括植物的次生代谢与化学合成。植物来源具有栽培周期长、产量不高、不可再生等缺点,难以应用于工业化生产;化学合成需要大量的能源,成本高,且造成环境污染。目前国内已报道了一些利用微生物转化生产用于医药、农药等的次生代谢物质如竹红菌素、虾青素、白藜芦醇[1-3]。微生物转化具有高效、高产、高强度、地成本,且不受季节、气候、地域等自然条件的限制,因此具有植物生产与化学合成不可比拟的优势,极具开发潜力和应用价值。
在植物苯丙氨酸代谢途径中,苯丙氨酸解氨酶(PAL)是苯丙氨酸代谢的第一步反应的酶与关键酶,PAL通过非氧化脱氨将L-苯丙氨酸(L-Phe)转化为肉桂酸,再经肉桂酸-4-羟化酶(C4H)转化为香豆酸,再由4-香豆酰-CoA-联结酶(4CL)、查尔酮合成酶(CHS)等催化形成黄酮类化合物,或者经芪合酶(STS)催化为白藜芦醇等芪合物(见图1)。
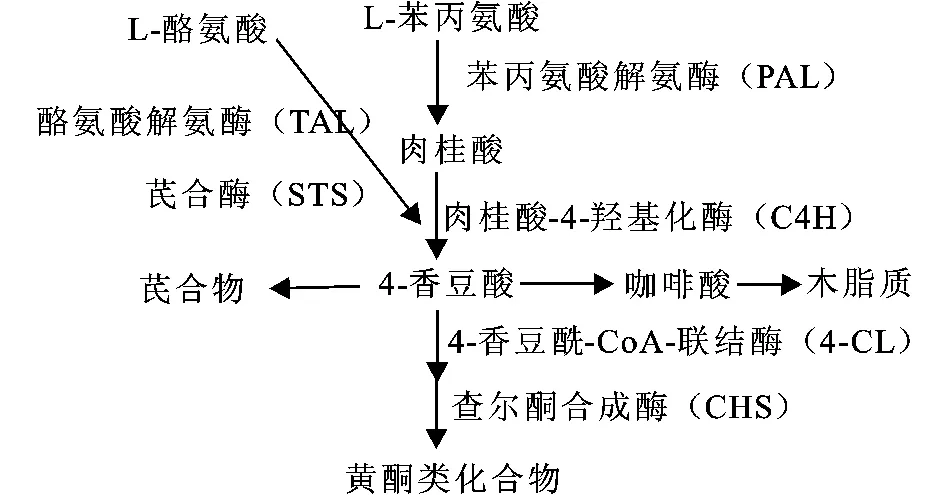
图1 植物苯丙氨酸代谢途径Fig.1 Metaolic pathway of phenylalaninie in plant
但目前已知的微生物中没有发现C4H和次生代谢物质的前体物质香豆酸[4],限制了由苯丙氨酸代谢途径生物转化生成代谢物质。TAL能不经C4H进行非氧化脱氨,直接将L-酪氨酸(L-Tyr)转化为香豆酸,因此研究人员对 TAL产生了极大的兴趣。目前发现的 TAL同时也具有PAL活性,已知植物 TAL对L-Tyr的亲和性都远低于L-Phe。近年来,在几种微生物中发现的 TAL对L-Tyr有较高的亲合力,经海藻酸钙固定 TAL基因重组的大肠杆菌,在125 L的生物反应器中以L-Tyr为底物进行转化,纯化到了12 kg的香豆酸[5],这为该酶的利用于微生物转化提供例证。
1 酪氨酸解氨酶的生物资源
目前发现 TAL存在于高粱、大麦、小麦、燕麦,水稻、玉米、甜甘蔗、欧芹等单子叶植物[6-7]。早在1968年,Bandoni等[8]报道了在担子菌中存在 TAL的活性。近年来,报道了不同微生物来源的 TAL的酶学性质。
2002年,Kyndt等[9]首次在荚膜红细菌(Rhodobacter capsulatus)分离到 TAL基因,基因表达于重组大肠杆菌中。纯化的TAL与L-Tyr和LPhe反应的最适p H值分别为8.5和9.4,与L-Tyr反应的转化量(Kcat)、催化效率(Kcat/Km)的值分别是L-Phe的2倍和150倍。
2006年,Kevin等[10]克隆到来源于(Rhodobacters sphaeroides)的 TAL基因,将该基因表达与大肠杆菌中,该酶与 Kyndt等[9]报道的Rhodobacter capsulatusTAL的氨基酸序列的同源性高达56%。经 SDS-PAGE纯化后,该酶分子量为55 000,与L-Tyr反应的最适p H值为 8.5~9.0,Kcat/Km是其与L-Phe反应的273倍,Kcat几乎相等。
2006年,Berner等[11]克隆到放线菌(S accharothrix espanaensis)的 TAL基因,在大肠杆菌中表达并纯化。该酶以L-Tyr为底物,p H值低于8.5时反应速率下降,p H值8.5以上时仍具有较高的反应速率,Km接近于 Kyndt等[9]报道的Rhodobacter capsulatuTAL,Kcat低于Rhodobacter capsulatuTAL。该酶以L-Tyr为底物的Kcat/Km是与L-Phe为底物的750倍,这个值是目前已报道最高的。
2007年,Vannelli等[12]从多种细菌和真核生物中分离出粘红酵母(Rhodotorula glutinis)的TAL,测定了该酶具有较高的胞内酶活力(0.014 3 U/mg蛋白)和最低的PAL/TAL活性比系数(1.68)。克隆该基因并表达于大肠杆菌中,纯化该酶的相对分子质量为287 000。
2007年,Vannelli等[13]测定9种真菌的以LTyr诱导的胞内 TAL活性,其存在于丝孢酵母(Trichosporoncutaneum)有最高的酶活性(0.179U/mg蛋白),但具有最低的 PAL/TAL活性比系数(0.8)。该酶纯化后研究表明,其相对分子质量为294 000的同源四聚体结构,亚基相对分子质量73 500,等电点为5.8,根据其N端和部分内部氨基酸序列,克隆了编码该酶的cDNA。cDNA序列分析表明,该序列于其他8种真菌的序列具有56%~62%的同源。
2007年,Xue等[14]测定4种微生物的以L-Tyr诱导的胞内 TAL活力,其中(Phanerochaete.chrysosporium)具有较高的酶活力(1.25U/g)。将TAL基因表达于大肠杆菌后用SDS-PAGE纯化,该酶相对分子质量为70 000,以L-Tyr为底物的Kcat/Km略高于以L-Phe为底物的Kcat/Km,PAL/TAL活性比系数为(0.7)。因该真菌能在较高温度下生长,据此推测该酶具有较好的热稳定性,研究表明在60℃高温下,3 h保持全酶的活力,4 h酶活力仍然可达72%。
2 酪氨酸解氨酶晶体结构研究
TAL与 HAL、PAL都属于芳香族解氨酶家族。2006年,Louie等[15]用 x射线探测了Rhodobacter sphaeroidesTAL(RsTAL)晶体的3级与 4级结构,其与以前报道的恶臭假单胞菌HAL、欧芹和酵母 PAL晶体结构类似。222对称点形成了同源四聚寡体,同源四聚体包含4个活性位置。3个不同单体参与构成活性位置的空腔,每个单体主要被5个自上而下螺旋状束有机围绕。束的肽链N端区域带3,5-二氢-5-次甲基-4H-咪唑-4-酮(MIO)辅酶,肽链另一端的C端部分构成了α螺旋外层,C端区域与中间的亚基相连并对同源四聚体起稳定作用和提供相连单体的活性空腔的外环。螺旋状束侧翼区连有凸出的发夹环,发夹环与第4和第5个螺旋束相连,侧翼区对单体与单体接口形成起作用。
RsTAL与恶臭假单胞菌 HAL缺少锲入到C端区域的附加区,欧芹和酵母PAL存在这个附加区,目前附加区的功能还不清楚。RsTAL与恶臭假单胞 HAL、真核的欧芹和酵母PAL多肽链的主链残基分别有34%、29%、30%一致。RsTAL与恶臭假单胞菌HAL的结构较真核PAL有更高相似,这与主链残基表现一致。
3 酪氨酸解氨酶的反应机制
TAL与 HAL、PAL都属于芳香族解氨酶家族,HAL和PAL的脱氨基催化反应有2种机制,一种被称为E1cB机制,底物的α氨基团与MIO的亚甲基碳直接发生亲核加成,另一种机制被称为Friedel–Crafts机制,富有电子的底物芳香环攻击引起了MIO的亚甲基碳电子的缺失[16]。
2006年,Louie等[15]用 X射线没能观察到RsTAL晶体中的L-Tyr转换,通过对RsTAL与香豆酸、咖啡酸、肉桂酸形成的复合体结构进行研究得到了以L-Tyr为底物的交联模型。在MIO亚甲基键长内,L-Tyr芳香碳环发生相当大平移,平移破坏α羧酸和羟基与 TAL形成的相互作用,这是Friedel– Crafts机制。另一方面,虽然在模型中L-Tyr的α氨基远离MIO辅酶,但丙烯酸基适度的构象变化将决定α氨基攻击MIO亚甲基碳的位置。如果α氨基攻击MIO亚甲基碳,氧空腔的水介导质子化和新形成的Asn203侧链酰胺的相互作用可能稳定MIO羰基氧的负电荷。通过对 TAL与AIP抑制剂形成的共价体进行观察,发现多肽链残基194到205的重排使Asn203明显接近MIO辅酶。但对突变体RsTAL H89F与AIP抑制剂的共价体进行结晶学观察,不足以支持了a氨基的直接攻击MIO亚甲基碳。因此这个模型基本上支持了Friedel– Crafts机制,但对 E1cB机制仍不能肯定。
4 酪氨酸解氨酶基因在次生代谢基因工程中的研究
目前已知的微生物中没有发现C4H和次生代谢的前体物质香豆酸[1],这限制了微生物通过苯丙氨酸代谢途径产生次生代谢物质。为了开辟次生代谢产物崭新的来源,科研人员将 TAL基因重组于微生物体内并成功获得了香豆酸、黄酮类、反式咖啡酸。
2007年,Vannelli等[12]将来源于Rhodotorula glutinisPAL/TAL基因整合在缺少C4HP-450基因及P-450还原酶且能产L-Phe的大肠杆菌中,实现了由 TAL途径将葡萄糖转化为香豆酸。2007年,Xue等[10]将来源于Rhodotorula glutinisTAL基因在大肠杆菌中表达,TAL将L-Tyr转化为香豆酸,该酶催化效率很低,但没有发现催化效率更高的TAL。抗高温的酶能通过提高温度而具有更高催化效率,随后发现了具有抗高温的Phanerochaete.chrysosporiumTAL,实验证明随温度的升高能产更高的香豆酸。
2004年,Watts等[17]将克隆于Rhodobacter sphaeroidesPAL/TAL基因与4CL基因、CHS基因共同表达于大肠杆菌中,测得柚皮素的含量高达20.8mg/L。2005年,Jiang等[18]首次将克隆于Rhodosporidium toruloidesTAL基因与4CL、CHS基因共同表达于啤酒酵母中,测得柚皮素含量为7mg/L,生松素含量为0.8mg/L,另外还获得了根皮素、2′,4′,6′-三羟基二氢查耳酮。
2006年,Berner等[11]将克隆于S accharothrix espanaensis的 TAL基因与 EC、CoA基因在大肠杆菌中共同表达获得了反式咖啡酸。
5 问题与展望
虽然TAL酶存在于微生物中,TAL基因在微生物次生代谢基因工程研究中已经取得一定成果,但目前该酶应用于微生物转化仍纯在一些问题,如酶活性、催化效率还有待于提高。随着进一步对TAL微生物资源的挖掘,酶学性质、结构,催化机制的深入研究,经过酶的结构修饰(如 KevinT等[10]对RsTAL突变体的研究发现,TAL对L-Phe和LTyr为底物的选择,单一的决定于His89,PAL获得该残基位点后也具有 TAL活性。)、基因重组、基因突变、酶的固定化、发酵工程的中游下游技术等的综合应用,TAL服务于微生物转化具有广阔的前景。
[1]梁晓辉,蔡宇杰,廖祥儒,等.竹黄发酵菌丝与竹黄子座成分比较分析[J].食品与生物技术学报,2009,28:705-708.LIANG Xiao-hui,CAI Yu-jie,LIAO Xiang-ru,et al.Analysis of the chemical compositions ofS hiraiasp.SUPERI-H168 fermented mycelia and the stromata of Shiraia bambusicola[J].Journal of Food Science and Biotechnology,2009,28:705-708-58.(in Chinese)
[2]胡建中,巩继贤,董庆霖,等.低氮促进红发夫酵母合成虾青素机理的研究[J].食品与生物技术学报,2009,28:91-96.HU Jian-zhong,GONGJi-xian,DONG Qing-Lin,et al.The mechanism of low nitrogen promoting astaxanthin biosynthesis in Phaff ia rhodozyma[J].Journal of Food Science and Biotechnology,2009,28:91-96.(in Chinese)
[3]唐永红,姚茂君,曹庸,等.一株产白藜芦醇真菌的分离及培养[J]食品与发酵工业,2009,7,33:30-33.TANG Yong-hong,YAO Mao-jun,CAO Yong,et al.Study on the isolation and culture of a resveratrol producing fungus[J].Food and Fermentation industries,2007,33:30-33.(in Chinese)
[4]Breinig S,Qi W,Sariaslani F,et a1.DNA and amino acid sequences of a tyrosine-inducible tyrosine ammonia lyase from Trichosporon cutaneum[P].美国专利:P6951751,2005.
[5]Trotman R,Camp C,Ben-Bassat A,et a1.Calcium alginatebead immobilization of cells containing tyrosine ammonia lyase activity for use in the production of p-hydroxycinnamic acid[J].Biotechnol Prog,2007,23:638-644.
[6]Neish A.Formation of m-and p-coumaric acids by enzymatic desamination of the corresponding isomers of tyrosine[J].Phytochem,1961,1:1-24.
[7]Appert C,Logemann E,Hahlbrock K,et a1.Structural and catalytic properties of the four phenylalanine ammonia-lyase isoenzymes from parsley(Petroselinum crispum Nym.)[J].Eur J Biochem,1994,225:491-499.
[8]Bandoni R,Moore K,Subba Rao P,et al.Phenylalanine and tyrosine ammonia-lyase activity in some basidiomycetes[J].Phytochemistry,1968,7:205-207.
[9]Kyndt J,Meyer T,Cusanovich M,et al.Characterization of a bacterial tyrosine ammonia lyase,a biosynthetic enzyme for the photoactive yellow protein[J].FEBS Lett,2002,512:240-244.
[10]Watts K,MijtsB N,Lee P,et al.Discovery of a substrate selectivity switch in tyrosine ammonialyase,a member of the aromatic amino acid lyase family[J].Chem Biol,2006,13:1317-1326.
[11]Berner M,Krug D,Bihlmaier C,et al.Genes and enzymes involved in caffeic acid biosynthesisin the actinomycete Saccharothrix espanaensis[J].J.Bacteriol,2006,188:2666-2673.
[12]Vannelli T,Qi W,Sweigard J,et al.Production of p-hydroxycinnamic acid from glucose inSaccharomyces cerevisiaeandEscherichia coliby expression of heterologous genes from plants and fungi[J].Metab Eng,2007,9:142-151.
[13]Vannelli T,Xue Z,Breinig S,Qi W,et al.Functional expression in Escherichia coli of the tyrosine-inducible tyrosine ammonia-lyase enzyme from yeast Trichosporon cutaneum for production of p-hydroxycinnamic acid[J].Enzyme and Microbiol Technol,2007,41:413-422.
[14]Xue Z,Michael M,Keith C,et al.Improved production of p-hydroxycinnamic acid from tyrosineusing a novel thermostable phenylalanine/tyrosineammonia lyase enzyme[J].Enzyme and Microbiol Technol,2007,42:58-64.
[15]Louie G,Bowman M,Moffitt M,et al.Structural determinants and modulation of substrate specificity in phenylalaninetyrosine ammonia-lyases[J].Chem Biol,2006,13:1327-1338.
[16]Schroeder A,Kumaran S,Hicks L,et al.Contributions of conserved serine and tyrosine residues Tocatalysis,ligand binding,and cofactor processing in the active site of tyrosine ammonia lyase[J].Phytochem,2008,69:1496-1506.
[17]Watts K,Lee P,Schmidt-Dannert C.Exploring recombinantflavonoid biosynthesis in metabolically engineered Escherichia coli[J].Chembiochem,2004,5:500-507.
[18]Jiang H,Wood K,Morgan J.Metabolic engineering of the phenylpropanoid pathway inSaccharomyces cerevisiae[J].Appl Environ Microbiol,2005,71:2962-2969.
