干旱及复水对谷子苗期根系形态特征及叶片解剖结构的影响
2024-04-01秦岭陈二影杨延兵黎飞飞王艳珂张梦媛管延安
秦岭 陈二影 杨延兵 黎飞飞 王艳珂 张梦媛 管延安


摘要:本试验选用抗旱性强的谷子品种济谷16 和旱敏感谷子品种鲁谷1 号为材料,设置盆栽试验,于出苗15 d 时停止浇水进行干旱胁迫处理,并于9 d 后复水,研究不同抗旱性谷子品种叶片以及根系形态指标对干旱-复水的响应。 结果表明:(1)与CK 相比,干旱胁迫导致两个谷子品种的根分枝數、根尖数和根体积显著减少;复水后,济谷16 的根尖数显著增加(P<0.05),而鲁谷1 号的根尖数进一步显著降低。 济谷16 的根分枝数在复水后的增加幅度大于鲁谷1 号。 (2)干旱胁迫下,两个品种叶片泡状细胞面积缩小,叶片厚度下降;复水后济谷16 泡状细胞面积与CK 相当,而鲁谷1 号仍显著小于CK(P<0.05)。(3)干旱胁迫下济谷16 下表皮气孔密度增加,上表皮气孔密度减小,且均与CK 差异显著(P<0.05);复水后,上表皮气孔密度略有增加但仍低于CK,下表皮气孔密度降低至CK 水平。 干旱胁迫下鲁谷1 号上、下表皮气孔密度较CK 均显著下降(P<0.05);复水后,上表皮气孔密度继续降低,而下表皮气孔密度显著增加至略高于CK 水平。 干旱胁迫后济谷16 上、下表皮气孔长度无显著变化,复水后较CK 和干旱处理均显著增加;鲁谷1 号上表皮气孔长度显著减小(P<0.05),复水后仍继续减小,但较干旱处理差异不显著;鲁谷1 号下表皮气孔长度较CK 增加但差异不显著,复水后继续增长,显著高于CK。 综上所述,干旱胁迫后复水,抗旱品种济谷16 通过根分枝数和根尖数的增加以及叶片泡状细胞面积、气孔密度快速恢复,使其表现出更强的形态适应性和自我调节能力。
关键词:谷子;干旱胁迫;复水;根系形态特征;叶片解剖结构;气孔
中图分类号:S515文献标识号:A文章编号:1001-4942(2024)01-0050-08
近年来,频繁发生的异常气候正在加剧干旱的发生程度和频率[1] 。 干旱已成为限制植物生长发育、造成作物减产的最主要非生物胁迫[2] 。 干旱胁迫下植物通过改变生物量的分配、增加叶片厚度、改变根系形态、上调脯氨酸和可溶性糖等渗透调节物质含量以适应干旱缺水[3] 。 实际生产中,作物一般处于干湿交替环境中,因此作物对干旱的适应能力不仅表现在干旱胁迫过程中,还依赖于复水后其形态、生理等指标的恢复能力[4] 。 作物对复水的响应方式是在胁迫解除后存在短暂的快速生长,以补偿或者部分补偿干旱造成的生长损失[5] 。
因此,复水恢复能力是植物适应干旱环境的一个重要机制,同时也是评价植物抗旱性的关键指标。大量研究表明,干旱胁迫和复水条件下,植物在形态结构、生理生化方面会发生一系列的适应性变化,这其中包括地上部叶片和地下部根系的协同作用[6] 。 干旱胁迫下苜蓿根干重、表面积、体积、密度和根尖数都表现为下降趋势,复水后根系会出现一定的补偿性生长[7] 。 干旱后复水对燕麦根总长、体积及平均根直径等指标具有促进作用[8] 。 研究表明,同一作物不同品种在干旱-复水过程中抗旱、恢复能力存在差异[9-10] 。 干旱胁迫导致作物叶片生理结构被破坏,但抗旱品种的叶面积在复水后会得到更好的恢复[11] 。
谷子[Setaria italica (L.) Beauv.] 起源于中国,其抗旱性强、耐瘠薄,广泛分布在我国西北、华北、东北的干旱和半干旱地区,在长期驯化和人工培育过程中不断适应了干旱和半干旱气候区的生长环境[12] 。 谷子具有叶片气孔密度大且内陷等抗旱特性[13] ,并有基因组序列小(约510 Mb)、自花授粉、生育期短、种质资源丰富等优势,使谷子成为研究非生物胁迫机制的理想模式作物[14] 。
目前研究者已对谷子抗旱生理机制[15] 以及干旱条件下形态指标[16] 、农艺性状[17] 等的变化进行了研究,但未从根系形态特征、叶片解剖结构等方面研究谷子对干旱的适应性以及复水后的恢复能力。 本研究采用盆栽控水法模拟土壤持续干旱胁迫及复水,研究不同抗旱性谷子品种在干旱和复水条件下根系形态、叶片解剖结构以及气孔特征的变化,旨在为谷子品种(系)抗旱性鉴定和优异抗旱种质创制提供科学参考。
1 材料与方法
1.1 试验材料
试验以抗旱性强的谷子品种济谷16、旱敏感谷子品种鲁谷1 号为材料。 种子均来源于山东省农业科学院作物研究所。
1.2 试验设计
试验于2019 年6 月在山东省农业科学院作物研究所旱棚内进行。 遮雨棚仅在下雨时使用,旱棚内光照和通风不受影响。 挑选籽粒饱满、大小一致的种子种植于黑色圆形塑料盆(高15 cm,口径11 cm,底径9 cm)内,盆内装250 g 育苗基质(有机质含量183.92 g/ kg,总氮1.12 g/ kg,总磷2.61 g/ kg,总钾0.59 g/ kg,pH 值5.69)。 每盆种10 粒,四叶期定苗至每盆3 株。 试验设3 个处理,其中,对照(CK) 保持土壤相对含水量(70±5)%;干旱处理(D) 在播种时土壤相对含水量70%,幼苗长至四叶一心时(出苗15 d) 停止浇水,使土壤水分持续下降至谷子幼苗叶片萎蔫(此时干旱处理9 d,土壤含水量约为对照的25%);复水处理(R)在停止浇水9 d 后进行,灌水至土壤相对含水量达(70±5)%。 干旱胁迫9 d以及复水24 h 后取样(图1),每处理各取3 盆(3次重复)。
1.3 测定项目及方法
1.3.1 根系形态指标的测定 将植株根系整体取出并在水桶内清洗干净,每个处理取3 株谷子幼苗根系,用WinRHIZO 植物根系分析测定仪( Win ̄RHIZO 2008a, Régent Instruments Inc.,Québec, Canada)扫描根系,测定根长度、根体积、根表面积、根直径、根尖数和根分枝数。
1.3.2 叶片显微结构 各处理分别取3 株谷子最新展开叶的中间部位,切成0.5 cm×1.0 cm 的小段,迅速放入装有FAA 固定液的5 mL 离心管中,置于冰盒中固定24 h。 将组织从固定液中取出,修切平整后置于脱水盒内进行脱水、浸蜡、包埋、切片、脱蜡、染色与封片等。 显微镜下观察切片并拍照,采用CaseViewer 软件(3DHIStech Ltd.Hungary)分别测定其上下表皮厚度、叶片厚度及泡状细胞面积,观察20 个视野取平均值。
1.3.3 叶表皮扫描电镜观测 分别切取不同处理植株新展开叶中部0.2 cm×0.2 cm 大小,冲洗干净后用3% ~4%戊二醛固定,4 ℃ 冰箱内固定12 h 后用磷酸缓冲液漂洗,进行丙酮梯度脱水、乙酸乙酯渗透、CO2干燥,然后用双面胶将叶片的上、下表面固定在样品台上,喷镀后拍照观察[18] 。
2 结果与分析
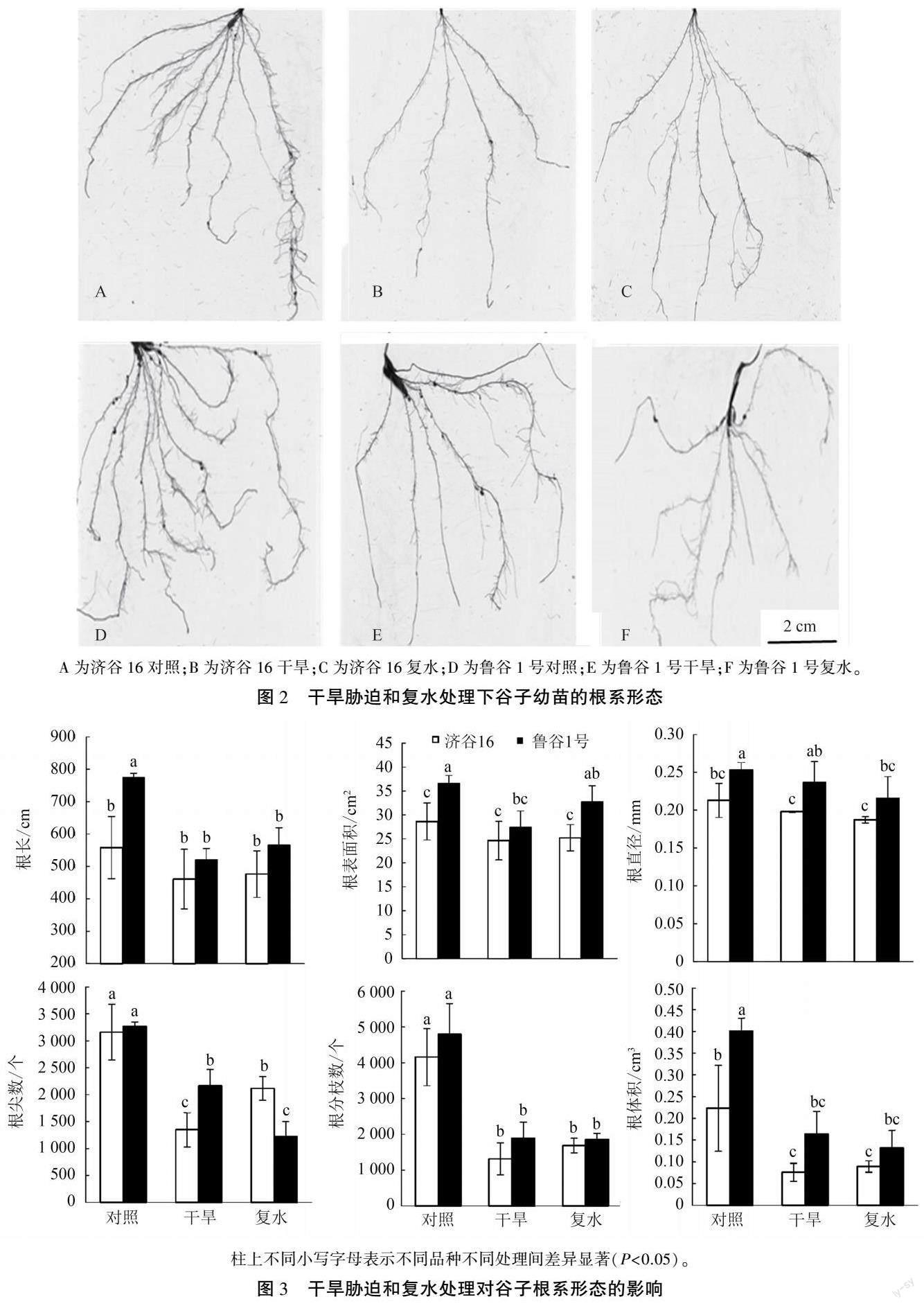
2.1 干旱胁迫和复水处理对谷子幼苗根系形态的影响
干旱处理抑制谷子幼苗根系发育(图2、图3)。 干旱处理对抗旱品种济谷16 的根长、根直径及根表面积影响较小,与CK 相比差异不显著,对其他根指标的影响均达显著水平。 干旱处理下,旱敏感品种鲁谷1 号除根直径较CK 无显著差异外,其他根部性状指标较CK 均显著减少。
干旱处理对谷子根分枝数、根尖数及根体积影响较严重,其中济谷16 和鲁谷1 号的根分枝数较对照分别减少68.38%和60.47%,复水24 h 后济谷16 的根分枝数较干旱处理增加28.43%,而鲁谷1 号的根分枝数在复水后未出现有效的补偿性增加。 干旱处理下济谷16 和鲁谷1 号的根尖数较对照分别减少57.33%和33.85%,复水24 h后济谷16 的根尖数较干旱处理增加56.82%,差异显著,而鲁谷1 号根尖数仍显著低于干旱处理。
干旱处理下济谷16 和鲁谷1 号的根体积较对照分别减少66.12%和59.10%,复水24 h 后济谷16根体积较干旱处理增加17.62%,而鲁谷1 号根体积并未出现补偿性增加。 可见,济谷16 的某些根部性状在干旱后下降比例较鲁谷1 号高,但其复水后的恢复能力也远高于鲁谷1 号。
2.2 干旱胁迫和复水处理对谷子幼苗叶片解剖结构的影响
谷子是C4植物,具有典型的Kranz 结构———“花环结构”,同时,巨大的泡状细胞能够充分吸水,对谷子抵御干旱非常有利(图4)。 由图4A 看出,供水充足条件下抗旱品种济谷16 幼苗叶片细胞结构清晰,排列较紧密,细胞间隙小且泡状细胞接近圆形;干旱处理后泡状细胞失水造成细胞上窄下宽,引起叶片轻微卷曲,降低蒸腾作用,从而减少干旱对细胞的损伤(图4B);复水24 h 后泡状细胞吸水叶片逐渐恢复平展(图4C)。 由图4D 看出,正常生长的鲁谷1 号幼苗叶片细胞结构清晰,泡状细胞较大;干旱胁迫处理后泡状细胞失水凹陷变形,细胞间隙变大,引起叶片卷曲闭合(图4E),复水24h 后泡状细胞吸水体积变大(图4F)。
由表1 可知,对照条件下,两个谷子品种幼苗叶片厚度差异不显著;干旱胁迫处理下,济谷16幼苗叶片厚度较对照减小11.18%,差异不显著,鲁谷1 号较对照减小13.65%,差异显著;复水24 h后,济谷16 基本恢复至对照水平,与对照差异不显著,鲁谷1 号仅略有恢复,仍显著低于对照。 两个谷子品种幼苗叶片上、下表皮厚度及泡状细胞面积在干旱胁迫后均比对照显著减小,其中济谷16 叶片泡状细胞面积较对照减少24.54%,鲁谷1号较对照减少43.04%。 复水后济谷16 叶片泡状细胞面积与对照相当,差异不显著,而鲁谷1 号仍显著小于对照。
2.3 干旱胁迫和复水处理对谷子幼苗叶片气孔特征的影响
济谷16 叶片上、下表皮气孔密度在正常供水条件(CK)下分别为175.83、152.83 个/ mm2,干旱胁迫后上表皮气孔密度降低30.28%(P<0.05),而下表皮气孔密度增加21.41%(P<0.05);复水后上表皮气孔密度略有增加但仍低于CK,下表皮气孔密度降低至CK 水平,差异不显著。 鲁谷1 号叶片上、下表皮气孔密度在CK 条件下分别为333.60、336.43 个/ mm2, 分别比济谷16 上、下表皮高89.73%、120.13%;受到干旱胁迫后其上、下表皮气孔密度均下降,分别降低24.47%、33.69% (P <0.05);复水后下表皮气孔密度恢复至与CK 无显著差异水平,而上表皮气孔密度未出现恢复性增加。
对照条件下两个品种叶片气孔长度存在差异:鲁谷1 号上、下表皮气孔长度差异显著,而济谷16 叶片上、下表皮气孔长度差异不显著。 干旱胁迫后济谷16 上、下表皮气孔长度较CK 分别增长15.71%、7.30%,但差异不显著;复水后均显著大于CK 和干旱处理。 干旱胁迫后鲁谷1 号上表皮气孔长度显著减小,较CK 减小58.94%,下表皮气孔长度增加14.92%,差异不显著;复水后上表皮气孔长度继续减小,下表皮气孔长度继续增大,但与干旱处理间差异不显著。
3 讨论
在受到干旱胁迫时,植物根系能够最早感知土壤缺水并做出适应性反应。 干旱环境下植物通过根系伸长、根直径减小等来提高根系的吸水性以确保其自身生长[19] 。 水稻在受到干旱胁迫时,单株次生根数、根系干重、表层根系干重等发生显著变化[20] 。 周广生等[21] 研究表明,不同水稻品种在不同干旱胁迫的不同时期根部特征存在差异,植株受到轻度以及中度干旱时根数量增加变慢,而受到严重干旱时根数量增加受到抑制,抗旱性强的品种根体积及干重降幅较小。 但也有研究发现相反的趨势,旱敏感水稻品种的根数量受到干旱胁迫后反而增加,而且多于抗旱品种,但是新生成的根都为小的细根,其茁壮程度远不如抗旱品种[22] 。 小麦根系特征与抗旱性也密切相关,干旱胁迫下抗旱品种通过增加最大根长来抵御干旱胁迫,并且根质量、总根长、根系总表面积、根系总体积等指标降幅显著小于旱敏感品种[23] 。 本研究中谷子受到干旱胁迫后根长、根表面积、根体积、根尖数等指标下降,这与前人研究结果一致[24] 。 复水后,济谷16 根尖数及根分枝数增加幅度大于鲁谷1 号,说明抗旱谷子品种根系补偿作用强于旱敏感品种,可以有效弥补前期干旱造成的损伤,这是其抗旱性强的原因之一。
葉片是高等植物光合作用的中心。 遭受干旱胁迫时,植物叶片的表型、生理、解剖结构等均能及时响应。 叶片的显微结构特征是植物适应干旱的一个重要方面,缺水时植物体内水分下降,造成叶片卷曲、萎蔫变黄。 禾本科作物叶片具有泡状细胞,它们在植物叶片卷曲中起到至关重要的作用,尤其与植物的抗逆性密切相关[25] 。 叶片厚度经常被作为衡量植物抗旱能力的一个重要指标[26] ,叶片越厚,其储水能力越强,有利于防止水分过度蒸腾[27] 。 抗旱糜子品种有较厚叶片、较大的叶肉表面积和泡状细胞长度等特点,增强了储水能力,降低了蒸腾,提高了光合效率,表现出较强的抗旱性[18] 。 不同植物对干旱胁迫的响应不同,大部分植物叶片厚度、表皮细胞厚度会在受到干旱胁迫时减小。 本研究表明,正常水分供应条件下不同抗旱性谷子品种叶片特征指标(叶片厚度、泡状细胞面积及上、下表皮厚度)并无显著差异。 泡状细胞失水少是抗旱性品种耐干旱环境的主要原因之一,使其细胞受伤害的程度小于旱敏感品种。
气孔是植物叶片与周围环境交换气体的器官,气孔密度越大,对CO2 的吸收量就越多,进而提高光合速率。 因此,干旱条件下,叶片气孔密度越大,越有利于植物短时间内充分利用有效CO2进行光合作用[28] 。 研究表明气孔的大小和数量跟植物蒸腾失水量直接相关,叶片表皮气孔在缺水条件下关闭而减少蒸腾作用;但有些植物对干旱的适应性有一定的限度,在严重缺水时,为了保证蒸腾作用气孔开度会增大[29] 。 同时,气孔密度大还有助于散热,从而避免高温引起叶绿体和原生质体发生变性[30] 。 许多旱生植物在干旱条件下通过使叶表皮气孔下陷、加大气孔密度或缩小孔径来应对缺水环境[31] ;但也有研究表明气孔密度越小则其抗旱能力越强,这是由于气孔密度与气体交换阻力呈负相关,当气孔密度减小时,气体交换阻力增大,蒸腾减少,因而增强抗旱能力[32] 。对油菜表皮细胞特征与干旱胁迫关系的研究也发现,抗旱性强的油菜品种气孔密度明显小于干旱敏感品种[33] 。 张盼盼等[18] 对不同糜子品种叶片解剖结构的研究发现,上、下表皮均有气孔分布,且下表皮气孔下陷,气孔密度大于上表皮。 一般认为叶片下表皮气孔数量高于上表皮有助于减少蒸腾作用造成的失水,进而提高植物适应干旱环境的能力。 而郭昱[34] 对362 份谷子核心种质叶片气孔特性的研究表明,谷子上、下表皮的气孔密度差异不显著。 本研究也发现两个品种叶片上、下表皮气孔密度差异较小,抗旱性强的品种济谷16 的上、下表皮气孔密度小于旱敏感品种鲁谷1号。 干旱胁迫处理下,两个品种的上、下表皮气孔密度均发生变化,但变化趋势不一致:济谷16 下表皮气孔密度增大,上表皮气孔密度减小;鲁谷1号上、下表皮气孔密度均出现下降趋势。 表明不同品种气孔密度和大小对干旱胁迫做出的响应不同。 抗旱性强的谷子品种可能存在一种合理的气孔调节机制,通过调节叶片上、下表皮气孔的开放和闭合来避免植株在干旱环境下因过度失水而死亡。
4 结论
干旱以及复水对不同抗旱性谷子品种叶片解剖结构、根系形态、气孔特性的影响不同。 抗旱谷子品种济谷16 干旱胁迫处理下幼苗叶片泡状细胞面积显著减小,根系各指标均呈现下降趋势。复水后,济谷16 幼苗根尖数较干旱处理显著增加,而鲁谷1 号未见有效补偿性增加;济谷16 的根分枝数有明显增加,但鲁谷1 号未出现补偿性增加。 干旱胁迫处理下,济谷16 幼苗叶片下表皮气孔密度显著增加,上表皮气孔密度显著降低,鲁谷1 号上、下表皮气孔密度均显著下降;济谷16叶片上、下表皮气孔长度无显著变化,鲁谷1 号上表皮气孔长度显著减小。 综上,干旱胁迫下,抗旱品种济谷16 的幼苗表现出更强的形态适应性、自我调节能力及复水后恢复能力。
参 考 文 献:
[1] Dai A. Increasing drought under global warming in observationsand models[J]. Nature Climate Change, 2013,3(1): 52-58.
[2] Trenberth K E, Dai A G, van der Schrier G, et al. Global war ̄ming and changes in drought [ J]. Nature Climate Change,2014,4(1): 17-22.
[3] Chaves M M, Flexas J, Pinheiro C. Photosynthesis under droughtand salt stress: regulation mechanisms from whole plant to cell[J]. Annals of Botany, 2009,103(4): 551-560.
[4] 黄海霞,连转红,王亮,等. 裸果木渗透调节物质和抗氧化酶活性对干旱的响应[J]. 干旱区研究,2020,37(1): 227-235.
[5] Liu C G, Wang Y J, Pan K W, et al. Photosynthetic carbonand nitrogen metabolism and the relationship between their me ̄tabolites and lipid peroxidation in dwarf bamboo (Fargesia rufaYi) during drought and subsequent recovery[J]. Trees: Struc ̄ture and Function, 2015,29(6): 1633-1647.
[6] Save R, Domingo R,Ruizsanchez M C,et al. Some physiologi ̄cal and morphological characteristics of citrus plants for droughtresistance[J]. Plant Science,1995,110(2): 167-172.
[7] 李振松,万里强,李硕,等. 苜蓿根系构型及生理特性对干旱复水的响应[J]. 草业学报, 2021,30(1): 189-196.
[8] 王晓雪,李越,张斌,等. 干旱胁迫及复水对燕麦根系生长及生理特性的影响[J]. 草地学报,2020,28(6): 1588-1596.
[9] 刘球,吴际友,李志辉. 干旱胁迫对植物叶片解剖结构影响研究进展[J]. 湖南林业科技,2015,42(3): 101-104.
[10] 朱维琴,吴良欢,陶勤南. 作物根系对干旱胁迫逆境的适应性研究进展[J]. 土壤与环境,2002,11(4): 430-433.
[11] 唐建,冯娟,刘云飞. 作物干旱复水补偿效应研究[J]. 湖北农业科学, 2021,60(1): 10-13,16.
[12] Doust A, Diao X M. Genetics and genomics of Setaria[M].Berlin: Springer, 2017: 61-72.
[13] 范光宇,赵治海,袁进成,等. 张杂谷3 号的解剖学结构与生物学特性的关系[J]. 华北农学报, 2014,29(增刊): 114-119.
[14] Muthamilarasan M, Prasad M. Advances in Setaria genomicsfor genetic improvement of cereals and bioenergy grasses[J].Theoretical and Applied Genetics, 2015,128(1): 1-14.
[15] Wang Y Q, Li L, Tang S, et al. Combined small RNA and de ̄gradome sequencing to identify miRNAs and their targets in re ̄sponse to drought in foxtail millet[J]. BMC Genetics, 2016,17(1): 57.
[16] 張文英,智慧,柳斌辉,等. 谷子全生育期抗旱性鉴定及抗旱指标筛选[J]. 植物遗传资源学报,2010,11(5): 560-565.
[17] 刘鑫,李会霞,田岗,等. 全生育期水分控制对谷子生长发育及品质的影响[J]. 作物杂志,2021(5): 181-186.
[18] 张盼盼,慕芳,宋慧,等. 糜子叶片解剖结构与其抗旱性关联研究[J]. 农业机械学报,2013,44(5): 119-126.
[19] 陈鹏狮,纪瑞鹏,谢艳兵,等. 东北春玉米不同发育期干旱胁迫对根系生长的影响[J]. 干旱地区农业研究,2018,36(1): 156-163.
[20] 李华,张玉屏,杨艳萍,等. 土壤水分状况对水稻表层根系和产量的影响[J]. 干旱地区农业研究,2006,24(1): 96-99.
[21] 周广生,徐才国,靳德明,等. 分蘖期节水处理对水稻生物学特性的影响[J]. 中国农业科学,2005,38(9): 1767-1773.
[22] 马廷臣,余蓉蓉,陈荣军,等. PEG6000 模拟干旱对水稻幼苗期根系的影响[J]. 中国生态农业学报,2010,18(6):1206-1211.
[23] 龚璞,李宗震,岳慧芳,等. 不同小麦抗旱品种种子萌发的根系特征及其与主要激素含量的关系[J]. 河南农业大学学报,2018,52(1): 16-21.
[24] 严四英,翁白莎,景兰舒,等. 干旱及旱后复水对夏玉米根系生长的影响[J]. 节水灌溉,2022(3): 75-81,91.
[25] 岑湘涛,沈伟,牛俊乐,等. 基于植物叶片解剖结构的抗逆性评价研究进展[J]. 北方园艺,2021(18): 140-147.
[26] 李广毅, 高国雄. 三种灌木植物形态特征及解剖结构的对比观察[J]. 水土保持研究,1995,2(2): 141-145.
[27] Dong X J, Zhang X S. Some observations of the adaptations ofsandy shrubs to the arid environment in the Mu Us Sandland:leaf water relations and anatomic features[J]. Journal of AridEnvironments, 2001,48(1): 41-48.
[28] 曹娟云,刘世鹏,白重炎,等. 干旱胁迫条件下狗头枣叶片解剖学结构比较研究[J]. 安徽农业科学,2008,36(8):3310-3311.
[29] 乔滨杰,王德秋,高海燕,等. 干旱胁迫下杨树无性系苗期光合与气孔形态变异研究[J]. 植物研究,2020,40(2):177-188.
[30] 孟庆杰,王光全,董绍锋,等. 桃叶片组织解剖结构特征与其抗旱性关系的研究[J]. 干旱地区农业研究,2004,22(3): 123-126.
[31] 杨九艳,杨劼,杨明博,等. 鄂尔多斯高原锦鸡儿属植物叶表皮特征及生态适应性[J]. 植物生态学报,2005,29(6):961-967.
[32] 刘照斌,宁俊,吕建洲. S3307处理对草地早熟禾叶片气孔密度和开度的影响[J]. 沈阳农业大学学报,2009,40(4):488-490.
[33] 陈智勇. 甘蓝型油菜叶片生长对干旱胁迫的响应分析[D].福州:福建农林大学, 2020.
[34] 郭昱. 谷子核心种质叶片气孔、乳突密度全基因组关联分析[D]. 晋中:山西农业大学, 2020.
