不同光周期对湛江等鞭金藻生长、岩藻黄素含量和光合作用及相关基因表达的影响
2024-02-23宋奕珩龚一富刘博悦郦露群王何瑜
宋奕珩 龚一富, 刘博悦 郦露群 王何瑜
(1宁波大学海洋学院,浙江 宁波 315832;2宁波大学食品与药学学院,浙江 宁波 315832)
湛江等鞭金藻(Isochrysiszhanjiangensis)是一种生长繁殖快、容易人工培养的海洋微藻,湛江等鞭金藻可产生一种重要的海洋药物——岩藻黄素(fucoxanthin,Fx),其最大产量可达23.29 mg·L-1[1]。岩藻黄素是硅藻、金藻和褐藻等特有的光合色素,在光合过程中起着关键作用[2]。岩藻黄素富含羰基、羟基和羧基等含氧官能团,使其具有更强的抗氧化活性[3]。岩藻黄素还被认为在抗衰老、抗肥胖以及对高血压和糖尿病的治疗中有独特的功能[4-5]。但是,目前岩藻黄素的生产存在产量低、难以满足市场需求的问题。因此,如何提高岩藻黄素含量是当前研究热点。
光照作为一个复杂的生态因子,对微藻生长发育的影响最为重要。在培养中改变光周期已经成为微藻次生代谢产物生产中的常用策略。有研究表明,新月柱鞘藻(Cylindrothecaclosterium)岩藻黄素的合成量在光周期为18L∶6D 时达到最大值23.6 mg·g-1DW[6]。而假微型海链藻(Thalassiosirapseudonana)在8L∶16D的光周期下获得最大的岩藻黄素产量(1.18 mg·g-1DW)[7]。但光周期是否促进湛江等鞭金藻细胞生长和岩藻黄素含量,目前还未见相关报道,优化光周期对湛江等鞭金藻细胞生长和岩藻黄素积累以及实现岩藻黄素规模化生产具有重要价值。
岩藻黄素生物合成受多种代谢通路的影响。章丽等[8]发现三角褐指藻(Phaeodactylumtricornutum)中八氢番茄红素脱氢酶基因(pds)、番茄红素β-环化酶基因(lcyb)和玉米黄素环氧酶基因(zep)等基因与岩藻黄质积累具有相关性。岩藻黄素含量还与光合作用相关,三角褐指藻经光合促进因子光合诱导素(photosynthetic induction factor,PIF)和抑制剂二氯苯基二甲基脲(dichlorophenyl dimethyl urea,DCMU)分别处理后,岩藻黄素含量相应的提高和下降[9-10],说明光合作用直接影响岩藻黄素积累。Li等[11]发现,随着光照时间减少,假微型海链藻和细孔海链藻(Thalassiosirapunctigera)的Rubisco 酶大亚基含量提高,可能与rbcL基因表达有关。光周期是否调控湛江等鞭金藻岩藻黄素合成通路和光合作用相关基因的表达,目前还未见相关报道。本试验通过研究不同光周期对湛江等鞭金藻细胞生长、岩藻黄素含量、叶绿素荧光参数及相关基因表达的影响,优化促进湛江等鞭金藻细胞生长和岩藻黄素积累的最佳光周期,探明岩藻黄素积累的光合生理和基因表达调控机理,旨在为进一步提高岩藻黄素含量和探明岩藻黄素的诱导表达调控分子机理提供理论依据。
1 材料与方法
1.1 材料与试剂
湛江等鞭金藻(I.zhanjiangensis)藻种,宁波大学海洋学院植物资源开发与利用实验室。植物RNA提取Plant RNA Kit,OMEGA 公司(美国);反转录试剂盒HiScript Ⅱ Q RT SuperMix for qPCR (+gDNA wiper)和实时荧光定量聚合酶连锁反应(quantitative real-time PCR,RTq-PCR)试剂ChamQ Universal SYBR qPCR Master Mix,诺唯赞生物科技有限公司(南京);其他药品及试剂,上海国药集团化学有限公司。
1.2 仪器与设备
cytoFLEX流式细胞仪,贝克曼公司(美国);UV-5200分光光度计,上海元析仪器有限公司;AquaPen-C AP 110-C 叶绿素荧光仪,FluorCam 公司(捷克);LC96 实时荧光定量PCR仪,Roche公司(瑞士)。
1.3 试验方法
1.3.1 湛江等鞭金藻的培养及处理 湛江等鞭金藻使用灭菌海水进行培养,配方参照DB3302/T 162-2018《海洋微藻饵料规模化培养技术规范》[12]。藻种培养于光照12 h/黑暗12 h(12L∶12D)、光照强度50 μmol·m-2·s-1、温度25 ℃的培养箱中。将生长至对数期的藻种以初始浓度为680 nm 下吸光度=0.1(A680=0.1)接种于海水培养基中,分别于不同光周期条件(0L∶24D、6L∶18D、12L∶12D、18L∶6D、24L∶0D)下培养,每组设3个生物学重复。
1.3.2 湛江等鞭金藻生长曲线测定 取生长至对数期的湛江等鞭金藻30 mL,在4 ℃下以5 000 r·min-1离心10 min后弃上清液,用海水培养基将藻泥以2倍逐级稀释后,用cytoFLEX 流式细胞仪计数各稀释组中湛江等鞭金藻的细胞密度,同时用UV-5200 分光光度计测定其A680。以细胞密度为横坐标、A680为纵坐标绘制标准生长曲线并建立回归方程。从试验处理开始,每24 h取藻液测定A680,并根据标准曲线回归公式换算成细胞密度,绘制生长曲线。每组设3个生物学重复。
1.3.3 湛江等鞭金藻岩藻黄素和叶绿素含量的测定 在第6天时,每瓶取3 mL藻液样品,使用分光光度法测定叶绿素含量[13]。以5 000 r·min-1离心10 min,弃上清液,向沉淀中加入3 mL无水乙醇,混匀,在60 ℃下避光浸提2 h。提取液以5 000 r·min-1离心10 min,取上清液在分光光度计下测定445 nm 下的吸光度(A445),根据吸光度计算岩藻黄素产量和含量,公式参照徐润洁等[14]并做修改。每组设3个生物学重复。
式中,Fxc为岩藻黄素产量(μg·L-1);Fxh为岩藻黄素含量(μg·105cells-1);A445为提取液在445 nm 下的吸光度;N为稀释倍数;V1为提取液体积(mL);V2为样品体积;A′为岩藻黄素浓度为1 g·L-1时在1 cm 光径比色皿中的理论吸收值,即1 600;细胞数单位为105个细胞。
1.3.4 湛江等鞭金藻叶绿素荧光参数测定 在不同光周期处理后24、48 和72 h 分别取各组湛江等鞭金藻藻液3 mL,稀释到近似A680后暗适应15 min,使用AquaPen-C AP 110-C叶绿素荧光仪分别测定光系统Ⅱ(photosynthesis Ⅱ,PSⅡ)最大量子产率(photochemical effiency,Fm/Fv)、PSⅡ实际光量子产率[Y(Ⅱ)]、光化学淬灭系数(photochemical quenching,qP)和非光化学淬灭系数(non-photochemical quenching,NPQ)。根据仪器说明书,将测量光强(Flash pulse)和饱和光强(Super pulse)设置为10%强度,光化光强(Actinic pulse)设置为培养时的光照强度,即50 μmol·m-2·s-1。每组设3 个生物学重复。
1.3.5 湛江等鞭金藻岩藻黄素相关合成通路的基因表达分析 根据实验室测得的湛江等鞭金藻转录组数据选取岩藻黄素合成通路的关键酶基因(pds、lcyb、zep1、zep2、zep3)和光合作用相关基因(rbcL,psbA),利用Primer Primer 5.0 软件设计定量引物(表1)。取不同光周期处理24 h后的藻液使用OMEGA 植物RNA 提取试剂盒提取总RNA,每组设3 个生物学重复,然后用HiScript Ⅱ Q RT SuperMix for qPCR (+gDNA wiper)试剂盒反转录。实时荧光定量PCR使用ChamQ Universal SYBR qPCR Master Mix 试剂进行扩增,每组设3 个技术重复。反应体系为10 μL ChamQ Universal SYBR qPCR Master Mix (2×)、0.4 μL 上游引物、0.4 μL 下游引物、8.2 μL 超纯水、1 μL 模板,共计20 μL。反应程序为95 ℃预变性30 s,40轮循环反应(94 ℃ 10 s,60 ℃ 30 s)。熔解曲线采集程序95 ℃ 15 s,60 ℃ 60 s,95 ℃ 15 s。使用2-ΔΔCT法计算目的基因的相对表达量。

表1 实时荧光定量PCR引物序列Table 1 Sequences of primers for real-time quantitative RT-qPCR
1.4 数据统计与分析
使用Microsoft Excel 2016 进行数据处理,使用SPSS 26.0 软件进行显著性检验、使用皮尔森(Pearson)相关系数来分析不同光周期下岩藻黄素含量与总叶绿素(total chlorophyll,chlT)含量、叶绿素荧光参数、基因表达量之间的相关性并进行主成分分析,以P<0.05为差异显著(*),P<0.01为差异极显著(**)。
2 结果与分析
2.1 光周期对湛江等鞭金藻生长的影响
根据流式细胞仪计数得到的细胞数和分光光度计测得的吸光度A680绘制标准生长曲线(R2为0.998 2),得到回归公式(3):
式中,Y为细胞密度(105cells·mL-1);X为吸光值(A680)。
由图1可知,0L∶24D处理下湛江等鞭金藻细胞无法生长。随着光照时间的延长,湛江等鞭金藻细胞密度呈现先上升后下降的趋势,以光周期为18L∶6D时的细胞密度最大,最高达到了148×105cells·mL-1,且与6L∶18D相比差异极显著(P<0.01),说明18L∶6D是湛江等鞭金藻细胞生长最适合的光周期。
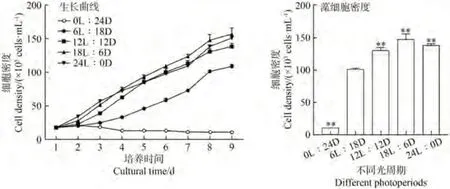
图1 光周期对湛江等鞭金藻生长趋势和藻细胞密度(第8天)的影响Fig.1 Effects of different photoperiod on cell growth trend and cell density(8th day) of I.zhanjiangensis
2.2 光周期对湛江等鞭金藻岩藻黄素含量的影响
研究不同光周期对湛江等鞭金藻岩藻黄素含量的影响,结果表明(图2-A),6L∶18D下岩藻黄素含量达到最高(0.041 μg·105cells-1)。相较于光周期为6L∶18D时,光周期12L∶12D、18L∶6D、24L∶0D 下岩藻黄素含量分别下降了5.3%、39.1%和63.5%(P<0.01),即随着周期中光照时间的延长,岩藻黄素含量呈下降趋势。岩藻黄素产量随周期中光照时间的延长呈先上升后下降趋势(图2-B)。与6L∶18D相比,12L∶12D下岩藻黄素产量极显著增加,达到了4.35 mg·L-1的最大值(P<0.01),随着光照时间的继续延长,岩藻黄素产量逐渐下降,在24L∶0D下达到1.62 mg·L-1的最小值(P<0.01),说明岩藻黄素产量受到细胞密度和单细胞岩藻黄素含量的共同影响。

图2 光周期对湛江等鞭金藻岩藻黄素含量和产量的影响Fig.2 Effect of photoperiod on the content and production of fucoxanthin in I.zhanjiangensis
2.3 光周期对湛江等鞭金藻叶绿素含量的影响
研究不同光周期对湛江等鞭金藻叶绿素含量的影响,结果表明(图3),叶绿素a(chlorophyll a,chla)和叶绿素c(chlorophyll c,chlc)含量均随光照时间的延长而下降。在光周期为6L∶18D时,总叶绿素(chlT)含量达到最高(0.033 μg·105cells-1),相较于光周期为6L∶18D时,光周期12L∶12D、18L∶6D、24L∶0D下的chlT含量分别下降21.7%、42.4%和60.1%。说明长时间光照不利于细胞积累叶绿素。根据相关性分析结果(表2),chlT含量与岩藻黄素含量之间呈显著正相关,相关系数为0.972。

图3 光周期对湛江等鞭金藻叶绿素含量的影响Fig.3 Effect of photoperiod on the production and content of chlorophyll in I.zhanjiangensis

表2 岩藻黄素含量与各指标间的相关系数Table 2 Correlation coefficient between fucoxanthin content and each index
2.4 光周期对湛江等鞭金藻叶绿素荧光参数的影响
研究不同光周期对湛江等鞭金藻叶绿素荧光参数的影响,结果表明(图4),随着处理时间延长,Fv/Fm在24L∶0D下第3天有极显著下降,其他组总体保持平稳。随着处理时间延长,Y(Ⅱ)和qP在6L∶18D下逐渐提高(P<0.01),在12L∶12D 和18L∶6D 下先上升后下降,在24L∶0D 下逐渐下降(P<0.01)。相关性分析结果显示(表2),Y(Ⅱ)和qP 与岩藻黄素含量间的相关性较强,相关系数分别为0.845和0.858。在所有组中都未测得非光化学荧光猝灭系数(non-photochemical quenching,NPQ),说明测定周期内所有处理未启动热耗散机制。
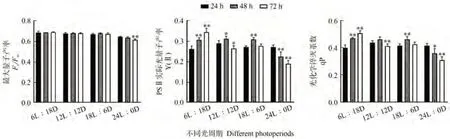
图4 光周期对湛江等鞭金藻叶绿素荧光参数的影响Fig.4 Effect of photoperiod on chlorophyll fluorescence parameters of I.zhanjiangensis
2.5 光周期对湛江等鞭金藻光合作用和岩藻黄素合成相关基因的影响
本试验选取了湛江等鞭金藻光合作用和岩藻黄素合成途径中较关键的一些基因,利用RTq-PCR 检测了光周期对这些基因相对表达量的影响,结果表明(图5),psbA和zep2基因相对表达量随着周期中光照时间的延长呈先下降后上升趋势,pds、rbcL、zep1、zep3和lcyb基因相对表达量呈下降趋势,但幅度有所不同。相关性分析表明(表2),zep家族的3个基因中,与zep1和zep2相比,zep3基因相对表达量与岩藻黄素含量的相关性系数最高,达到了0.863,为强相关性(P>0.8),说明zep3基因可能是响应光周期变化中最相关的zep基因。岩藻黄素含量与rbcL、pds、lcyb、zep3基因变化的相关系数都超过了0.7,存在较强的相关性,说明岩藻黄素的变化是基因表达水平改变的结果。

图5 不同光周期下湛江等鞭金藻各基因转录的差异Fig.5 Differences in gene transcription of I.zhanjiangensis under different photoperiod
2.6 光周期对湛江等鞭金藻各参数的主成分分析结果
主成分分析结果表明(图6),所有的参数被降维到了两个主成分中(RC1、RC2),其中,RC1为叶绿素荧光参数,贡献率为51.54%,RC2 为光合作用和岩藻黄素合成通路相关基因,贡献率为42.77%,累计贡献率为94.31%。主成分分析表明,光合作用和岩藻黄素合成相关基因的变化是影响岩藻黄素含量变化的主要因素。光合生理参数中的chlT 含量、Y(Ⅱ)和qP 与岩藻黄素含量(Fx)距离较接近,说明岩藻黄素含量的变化受到光合作用的影响。结合相关性分析结果,说明岩藻黄素合成受到光合作用和基因表达的共同影响,且rbcL、pds、lcyb和zep3基因是影响岩藻黄素合成的主要基因。

图6 湛江等鞭金藻各参数的主成分分析结果Fig.6 Results of principal component analysis of parameters of I.zhanjiangensis
3 讨论
本研究发现,湛江等鞭金藻生长的最适光周期为18L∶6D,与等鞭金藻(Isochrysisgalbana)、微拟球藻(Nannochloropsisgaditana)细胞生长的最适光周期相一致,但与布朗葡萄藻(Botryococcusbraunii)细胞(24L∶0D)和蛎甲藻属微藻(Ostreopsiscf.ovata)细胞生长的最适光周期(12L∶12D)不同[15-18],这可能和微藻的种类有关。微藻以在黑暗下进行分裂,在光照中进行生长的节律来提高光合效率[19],本试验发现24L∶0D 光周期下细胞密度降低,可能是因为这种节律被打破,导致藻细胞的光合效率降低,细胞密度下降。
岩藻黄素会与叶绿素a/c 和某些蛋白结合形成捕光天线复合体并向后传递能量,推动暗反应进行[20]。作为一种天线色素,岩藻黄素含量与光照条件密切相关。Wang 等[6]发现,在24∶0D 下会导致新月柱鞘藻(Cylindrothecaclosterium)的Rubisco 酶等蛋白及其转录调节因子表达下调,进而造成岩藻黄素含量减少,同时微藻会发生光抑制进而造成岩藻黄素含量降低。Palanisamy等[7]发现,假微型海链藻的岩藻黄素含量在8L∶16D 下获得最大值,并且随着光照时间从8 h 增加到24 h,岩藻黄素含量逐渐下降,与本研究趋势一致。而岩藻黄素产量受到细胞密度和单细胞岩藻黄素含量的共同影响,这种变化趋势是由光照时间导致的细胞密度变化和细胞内岩藻黄素含量的变化所致。湛江等鞭金藻在12L∶12D 下获得了最大的岩藻黄素产量,因此需要进一步研究岩藻黄素合成量和细胞生长之间的平衡点,从而在实际运用中达到最高的岩藻黄素产量。
本研究发现,叶绿素和岩藻黄素含量间存在较强的相关性,Nicklisch等[21]计算了不同藻中岩藻黄素含量与叶绿素含量的比值,发现光周期的长短对其影响很小,与本试验的结果一致。因此,当光照时间不足时,微藻可能为了提高光合效率,通过负反馈持续增加叶绿素含量,造成岩藻黄素含量提高。同时也有研究表明,过长的光照时间不利于叶绿素的产生[22-23],这可能和叶绿素合成相关酶受到的光调控有关。如植物中存在一些转录因子如HY5 等可在光照下促进基因的表达,而植物光敏色素互作因子PIFs 可在黑暗中抑制Chl 合成基因的表达[24]。Palanisamy 等[7]认为,在过长的光周期下,浮游植物细胞会减少叶绿素a 的合成和积累,从而改善过度光能引起的光损伤或光抑制,并且在24 h 光照下发生的光抑制现象也会造成叶绿素的降解[25]。
叶绿素荧光参数可以评价PSⅡ光能捕获、电子传递和光能损耗等过程,并且具有较高的灵敏性[26]。24L∶0D 下湛江等鞭金藻Fv/Fm下降,说明PSⅡ最大光能转化效率降低,植物发生了光抑制,这解释了岩藻黄素与叶绿素含量的下降[27]。当光抑制发生时会产生活性氧(reactive oxygen species,ROS),造成PSⅡ反应中心D1 蛋白破坏,但植物体存在损伤修复的机制,可以合成新的D1 蛋白保持PSⅡ功能[28],这解释了PsbA在24L∶0D 组中表达量的提高。6L∶18D 下Y(Ⅱ)和qP的逐渐提高,说明光合系统实际光能转化效率和电子传递活性显著增强,使微藻在缩短的光照时间内提高光合效率从而利用更多的光能[27]。而24 h 光照下Y(Ⅱ)和qP 的逐渐下降表明PSⅡ的反应中心受损。在本试验中未能测到NPQ 数据,表明在本试验条件下湛江等鞭金藻并未启动NPQ机制进行热耗散。
pds和lcyb基因在岩藻黄素合成中十分关键,pds编码的八氢番茄红素脱氢酶是岩藻黄素合成的限速酶[29],lcyb位于类胡萝卜素合成的分支点[30]。zep基因位于岩藻黄素合成末端的位置[12],对三角褐指藻的研究发现,3 个zep基因在藻中和不同因素影响下的表达情况不同[31-32],胺处理和尿素处理使zep2大量表达,蓝光处理使zep1和zep3表达水平上升,海带提取物会使zep3的表达水平发生最显著的上调。因此,3个zep基因在光周期下的不同变化趋势可能说明这3 个基因具有不同的作用,zep3基因与岩藻黄素含量的相关性系数最高,可能说明光周期通过影响zep3表达改变了岩藻黄素合成。rbcL基因编码暗反应关键酶Rubisco酶的大亚基[33],Minoda等[34]发现,黑暗环境会提高红藻(Cyanidioschyzon merolae)rbcL基因表达水平并在暗-光转换时激活光合作用,因此6L∶18D组中rbcL基因表达水平上调可能是为了提高光合作用。
4 结论
光周期通过对湛江等鞭金藻光合作用和岩藻黄素合成相关基因的共同作用,影响了藻体中岩藻黄素的合成和积累,不同基因受光周期影响的变化趋势不同,光周期可能主要通过影响zep3表达影响了岩藻黄素合成。通过改变光周期来提高岩藻黄素的含量,有利于岩藻黄素生产,但仍需解决细胞生长和岩藻黄素合成之间的平衡问题。
