特色香料作物油樟叶内生真菌多样性及其挥发性成分分析
2024-02-23刘雯雯胡连清杰周万海冯瑞章
刘雯雯 胡连清 陈 露 杨 丽 魏 琴 周 杰周万海 , 冯瑞章
(1宜宾学院,四川省油樟工程技术研究中心,四川 宜宾 644007;2宜宾学院,农林与食品工程学部,四川 宜宾 644007;3四川宸煜林业开发有限责任公司,四川 宜宾 644007)
油樟(Cinnamomumlongepaniculatum)是我国特有的木本香料林业资源,其中提取的天然芳香油在医药、日化和食品等行业发挥着重要作用[1]。油樟叶部含油量最高,主要成分为1,8-桉叶油素[2]、α-松油醇、β-蒎烯和γ-松油烯等活性成分[3-4],是医药、日化和食品等生产的重要原料[5],且有研究表明油樟精油具有抗氧化、抑菌和抗肿瘤等作用[6-7]。关于油樟精油的提取和成分分析已有相关报道,例如尹浩等[8]研究表明采用酶辅助同蒸馏萃取法提取的油樟叶精油收率最高;宁登文等[9]发现4、8 和12 月采集的油樟叶出油率最高,树龄21~40 年且嫩叶和混合叶的相对出油率最高;程贤等[10]研究结果显示春季油樟叶精油提取率高于秋季,但两季精油主要成分无显著差异。四川宜宾油樟叶含油量约为江西、广东及台湾等省油樟油量的2.5倍,且比香樟叶含油率高2 倍[11],推测与其共生微生物等特殊环境条件有关[12]。
植物内生真菌指长期或生活史某一个阶段定殖于健康植物内部而不引起宿主明显病理症状的微生物[13],研究证实其种类和数量随宿主植株的种类、季节或器官等不同而不同[14]。近年来,蛇足石杉[15]、粉色西番莲[16]、樟叶越橘[17]等不同生境植物内生真菌得到了广泛研究。部分内生菌可产生与宿主相同或相似的活性物质,在农业、医药和生物防治等方面具有较大潜力[18]。关于油樟内生菌的多样性和功能性研究已有一些报道,如冯瑞章等[19]从油樟中筛选到3 株具有溶磷作用的内生细菌,对植物生长、抗性和生物防治具有应用效果;黄金凤[20]通过紫外线诱变筛选出油樟叶中高产α-松油醇的内生细菌;严宽等[21-22]发现用油樟内生真菌2J1 和3J1 的孢子悬浮液处理油樟叶21 d 能显著提高叶片的挥发油含量,建立的青霉菌属油樟内生真菌和茉莉酸甲酯培养体系,促使了油樟细胞挥发油的合成。谭韵雅等[23]发现油樟内生真菌YY26 和YG42可促进1,8-桉叶油素等成分的积累,抑制了YG71代谢产物的分泌。但关于油樟叶可培养内生真菌的多样性、菌株与产油量关联性及菌株挥发性成分分析研究还较少。基于此,本研究以四川宜宾油樟为材料,测定不同株系和不同季节油樟叶的芳香油量,研究其叶部内生真菌的多样性和分布情况,探讨不同株系、季节的菌株种类与含油量关系,并测定菌株发酵液的挥发性成分,以期为油樟内生真菌资源多样性及提供可应用的功能菌株奠定基础。
1 材料与方法
1.1 试验材料
油樟叶采集于四川省宜宾市高县月江森林经营所油樟母本园(北纬28°47′77″,东经104°59′76″)的5 个株系(1、14、16、40 和53 号株系),根据区域气候特点,采样时间为3月下旬(春季)、6月下旬(夏季)、9月下旬(秋季)和12月上旬(冬季)。采样时,每个品系随机选取成年健康油樟3株,每株树间隔大于20 m,使用单绳攀爬等方式,到达大树顶端,用灭菌剪剪取东南西北4 个方向的树叶,每个方向取3条树枝,选取枝条上端健康完整的树叶,每个树枝取10片,混合放入4 ℃冰盒带回实验室,用于后续内生真菌分离;枝条剩余叶片全部收集后用于油樟油提取。马铃薯葡萄糖琼脂(potato dextrose agar,PDA)营养琼脂、麦芽提取物琼脂、察氏、马丁、卵磷脂吐温-80 营养琼脂、玉米粉琼脂均为成品培养基,购自北京三药科技有限公司。
1.2 试验方法
1.2.1 油樟叶精油的提取 将采集的部分油樟叶放置于室内避光处,待其自然干燥至恒质量后,根据《中华人民共和国药典 2020 年版 四部》[24]通则2204 挥发油的测定方法进行提取和测量。
1.2.2 油樟叶可培养内生真菌的分离纯化 收集健康新鲜的油樟叶于无菌取样袋中,自来水冲洗组织表面2 h,转入到灭菌瓶用无菌水冲洗3 次,95%乙醇漂洗1 min,再用无菌水冲洗1 次,4%(有效氯含量)次氯酸钠漂洗4 min,最后用95%乙醇漂洗1 min,无菌水冲洗5 次,每次30 s。设置漂洗对照、印记对照和环境对照[25]。吸干油樟叶表面水分,利用无菌剪刀将叶片剪成约4 mm2的小叶块,分别接种到PDA 营养琼脂、麦芽提取物琼脂、察氏、马丁、卵磷脂吐温-80 营养琼脂和玉米粉琼脂培养基上,每皿接种5 片,设置10 个重复,28 ℃恒温培养。每24 h 观察菌落情况并计数,根据各平板菌落形态随机挑选具有代表性的单菌落,连续传代培养至菌落形态单一。
1.2.3 内生真菌的内转录间隔区(internal transcribed spacer,ITS)序列分析 采用真菌通用引物(ITS1:5′-TCCGTAGGTGAACCTGCGG-3′、ITS4:5′-TCCTCCGC TTATTGATATGC-3′)扩增ITS 序列。PCR 反应体系为50 μL:ddH2O 21 μL,2×Taq PCR Master Mix 25 μL,20 μmol·L-1正向引物1 μL,20 μmol·L-1反向引物1 μL,DNA 模板2 μL。PCR 反应条件:94 ℃预变性10 min;94 ℃变性30 s,58 ℃退火30 s,72 ℃延伸45 s,35 个循环;72 ℃终延伸10 min。扩增产物用1%琼脂糖凝胶电泳检测,合格后送上海生工生物工程公司进行序列测定。将所得序列信息提交至NCBI 网站(https://www.ncbi.nlm.nih.gov/)BLAST程序进行序列同源性比对。
1.2.4 油樟叶内生真菌群落多样性分析 选用4 个指标评价油樟叶内生真菌的多样性,即物种丰富度S、Shannon-Weiner 多样性指数H′、Simpson 优势度指数D、均匀度指数E,具体计算公式为:
式中,N为样本全部属数之和;Pi为属相对频率,指样本中分离某属的菌株数占分离总菌株数的百分比。对油樟叶不同株系或季节产油率与叶中可培养内生真菌多样性指数之间的相关性进行Pearson 相关分析,探究产油率对油樟叶中可培养内生菌群落组成的影响。
1.2.5 油樟叶内生真菌挥发性成分分析 通过气相色谱/质谱联用技术(gas chromatograph/mass spectrograph,GC/MS)对内生真菌发酵液进行挥发性成分分析。GC条件为:HP-5毛细管气相色谱柱(30 m×320 μm×0.25 μm,安捷伦,美国),载气为高纯N2,流量26.56 mL·min-1,程序升温,起始温度80 ℃保持3 min,以5 ℃·min-1升温至90 ℃保持2 min,5 ℃·min-1升温至170 ℃保持2 min,5 ℃·min-1升温至220 ℃保持2 min 结束。进样口温度240 ℃,进样量1 μL,不分流进样,尾吹流量25 mL·min-1。MS条件为:电子轰击(electron impact,EI)离子源方式,离子源温度为200 ℃,离子化电压70 eV,扫描范围50~650m/z[26]。根据上述分析结果,比较筛选能产与宿主挥发油主要成分相同的菌株。
2 结果与分析
2.1 油樟叶提取的精油含量
由表1可知,选用的5株油樟株系叶产油率存在差异。春季和夏季5 个油樟株系叶产油率均表现为16>40>53>14>1,但秋季表现为53>16>1>14>40,冬季表现为14>16>53>40>1。由季节平均产油率结果可知,春季油樟叶产油率最高,夏季最低,但各指标差异均不显著。各株系油樟叶产油量以16号最高,1号最低,且差异显著(P<0.05),表明株系特征是决定油樟叶产油率的关键。
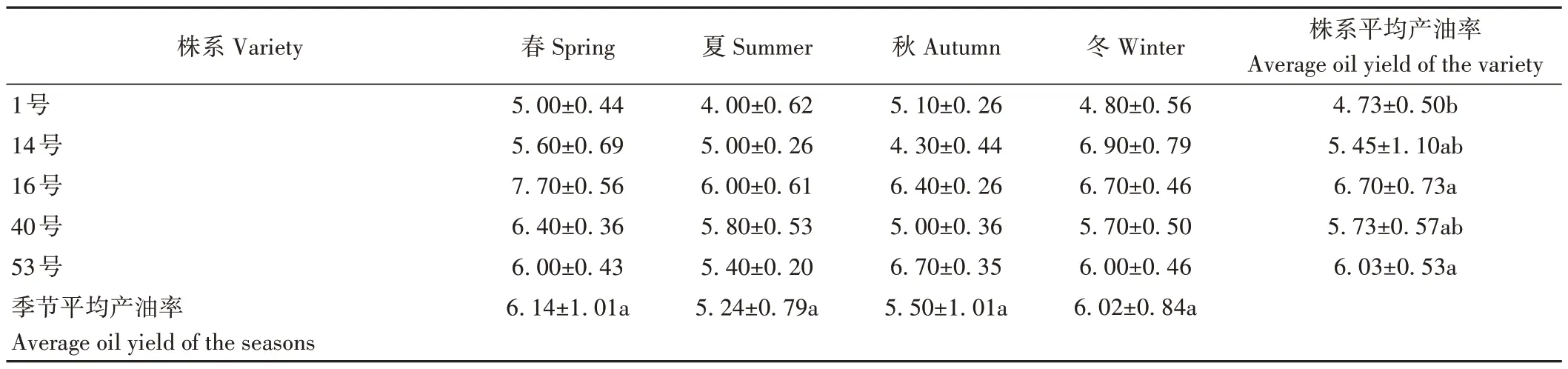
表1 不同株系和季节油樟叶的产油率Table 1 Oil production rate of C.longepaniculatum leaf in different varieties and seasons/%
2.2 油樟叶内生真菌的分离及群落组成分析
从油樟叶分离的菌株经过分离纯化共获得43 株内生真菌(电子附表1),不同株系和季节的内生真菌菌株数存在差异,根据株系分类,1、14、16、40、53 号油樟株系分别分离到8、12、8、10和5株菌株;根据季节分类,春夏秋冬季油樟叶分别分离到7、25、2和9株菌株。经鉴定43 株内生真菌归属于2 门、4 纲、9 目、11 科、13属,其中优势门为子囊菌门(Ascomycota),优势纲为座囊菌纲(Dothideomycetes),优势目为葡萄座腔菌目(Botryosphaeriales)、格孢腔菌目(Pleosporales),优势科为叶点霉科(Phyllostictaceae),优势属为叶点霉属(Phyllosticta)。不同株系油樟叶内生真菌分布显示,1 号株系油樟叶分离的8 株菌鉴定为1 门2 纲4 目4 科5 属,5 个属包括链格孢属(Alternaria)、斜锉蛤属(Pseudoplagiostoma)、刺盘孢属(Colletotrichum)、球座菌属(Guignardia)和Phyllosticta属;14号株系油樟叶分离的12 株菌鉴定为1 门3 纲5 目7 科7 属,7 个属包括Phyllosticta、Colletotrichum、拟茎点霉属(Phomopsis)、Pseudoplagiostoma、间座壳属(Diaporthe)、黑孢霉属(Nigrospora)和曲霉属(Aspergillus);16 号株系油樟叶分离的8 株菌鉴定为1 门3 纲5 目5 科6 属,6 个属包括枝孢菌属(Cladosporium)、Guignardia、Phyllosticta、Colletotrichum、Nigrospora和青霉菌属(Penicillium);40号株系的10株菌鉴定为1门3纲4目4科5属,5个属包括Aspergillus、Guignardia、Phyllosticta、Colletotrichum、Pseudoplagiostoma;53号株系油樟叶分离的5株菌鉴定为2 门4 纲5 目5 科5 属,5 个属包括Colletotrichum、Phyllosticta、Penicillium、蚁巢伞属(Termitomyces)和黑管菌属(Bjerkandera)。根据不同季节油樟叶内生真菌分离结果可知,春季7株菌鉴定为1门2纲3目5科5属,5 个属包括Cladosporium、Colletotrichum、Phomopsis、Pseudoplagiostoma和Diaporthe;夏季的25株菌鉴定为1门3 纲3 目3 科5 属,5 个属包括Guignardia、Phyllosticta、Aspergillus、Penicillium和Colletotrichum;秋季2 株菌鉴定为1 门2 纲2 目2 科2 属,分别为Alternaria和Phyllosticta;冬季9株菌鉴定为2门2纲5目5科5属,5个属包括Colletotrichum、Pseudoplagiostoma、Nigrospora、Termitomyces和Bjerkandera。
通过图1可知,5个不同株系油樟叶内生真菌属类数量差异较小,其中5 株共有属为Colletotrichum和Phyllosticta。Penicillium和Cladosporium为16 号株系特有属。表明油樟不同株系叶中可培养内生真菌具有一定相似性,但同时也具备各自多样性。

图1 基于属水平的不同株系油樟叶内生真菌群落图Fig.1 Community of endophytic fungi in different varieties of C.longepaniculatum leaves based on genus level
由图2可知,春夏冬3季属类数量相同,秋季最少,仅有2属。春夏秋冬季未见共有属,春夏秋3季共有属为Colletotrichum。Cladosporium、Phomopsis、Diaporthe为春季特有属;Guignardia、Aspergillus、Penicillium 仅出现在夏季;Alternaria仅出现在秋季;Nigrospora、Termitomyces、Bjerkandera仅出现在冬季,可能与不同季节产油率有关。表明季节亦是影响油樟不同株系叶内可培养内生真菌多样性的主要因素之一。
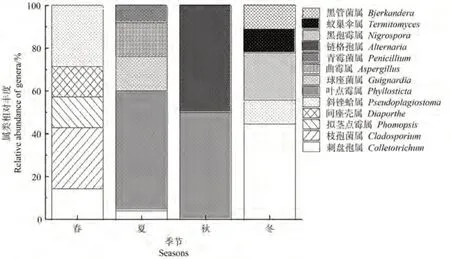
图2 基于属水平的不同季节油樟叶内生真菌群落图Fig.2 Community of endophytic fungi in different seasons of C.longepaniculatum leaves based on genus level
2.3 油樟叶内生真菌的多样性及相关分析
多样性分析可显示宿主植物在生长过程中内生菌群落结构的变化规律,常采用物种丰富度(S)、Shannon-Wienner 指数(H′)、Simpson 指数(D)和均匀度指数(E)等反映多样性指标。其中H′与物种丰富度关系最密切,数值越大表明物种数量越多,分布越均匀;D数值越大,显示优势种越明显;E敏感性高于H′,用于分析内生真菌群落分布的均匀程度。通过表2和3可知,不同株系油樟叶内生真菌S指数差异不明显,14 号株系最大,16号株系次之;H′指数表现为16号>53号>14号>40 号>1 号;D指数为16 号>53 号>40 号>1 号>14 号;均匀度指数E为53号株系最大,16号株系次之,14号株系最小。不同季节油樟叶内生真菌S指数春季和夏季值最大,秋季最小;H′指数和D指数均表现为春季>冬季>夏季>秋季;4 季均匀度指数E差异不明显,冬季最大,秋季最小。由此可知,16 号株系和春季油樟叶内生真菌群落的多样性相对最丰富,53 号和冬季次之,1号和秋季最低。因此,株系和季节是影响油樟叶中可培养内生真菌多样性的重要因素。进一步Pearson 相关分析结果发现,不同株系或不同季节油樟叶精油产率与叶中可培养内生真菌多样性指数无显著相关关系,挥发油对可培养内生真菌的影响还应做进一步分析。

表2 不同株系油樟叶内生真菌的多样性指数Table 2 Diversity index of endophyte fungi in C.longepaniculatum leaves of different varieties

表3 不同季节油樟叶内生真菌的多样性指数Table 3 Diversity index of endophyte fungi in C.longepaniculatum leaves of different reasons
2.4 产香能力测定
对43 株内生真菌发酵液的GC/MS 分析结果中(表4),共鉴定出69种主要挥发性成分,含醇类化合物13种、酸类化合物11种、酯类化合物11种、烯类化合物6 种、酮类化合物10种、醛类化合物7种、酚类化合物8种和其他类3 种。挥发性成分中有多种产香气物质,可作为食品添加剂和香料配方,同时也检测到部分菌株产与油樟精油主要成分相同的物质,如X29 内生真菌发酵液产生的桉叶油醇,相对质量分数为0.39%;D72 发酵液产生的芳樟醇,相对质量分数为0.42%;A39 发酵液产生的红没药醇,相对质量分数为0.97%(表5)。X29和A39均属于Phyllosticta。

表4 油樟叶内生真菌挥发性成分鉴定结果Table 4 Results of identified volatile components of endophyte fungi from C.longepaniculatum leaves
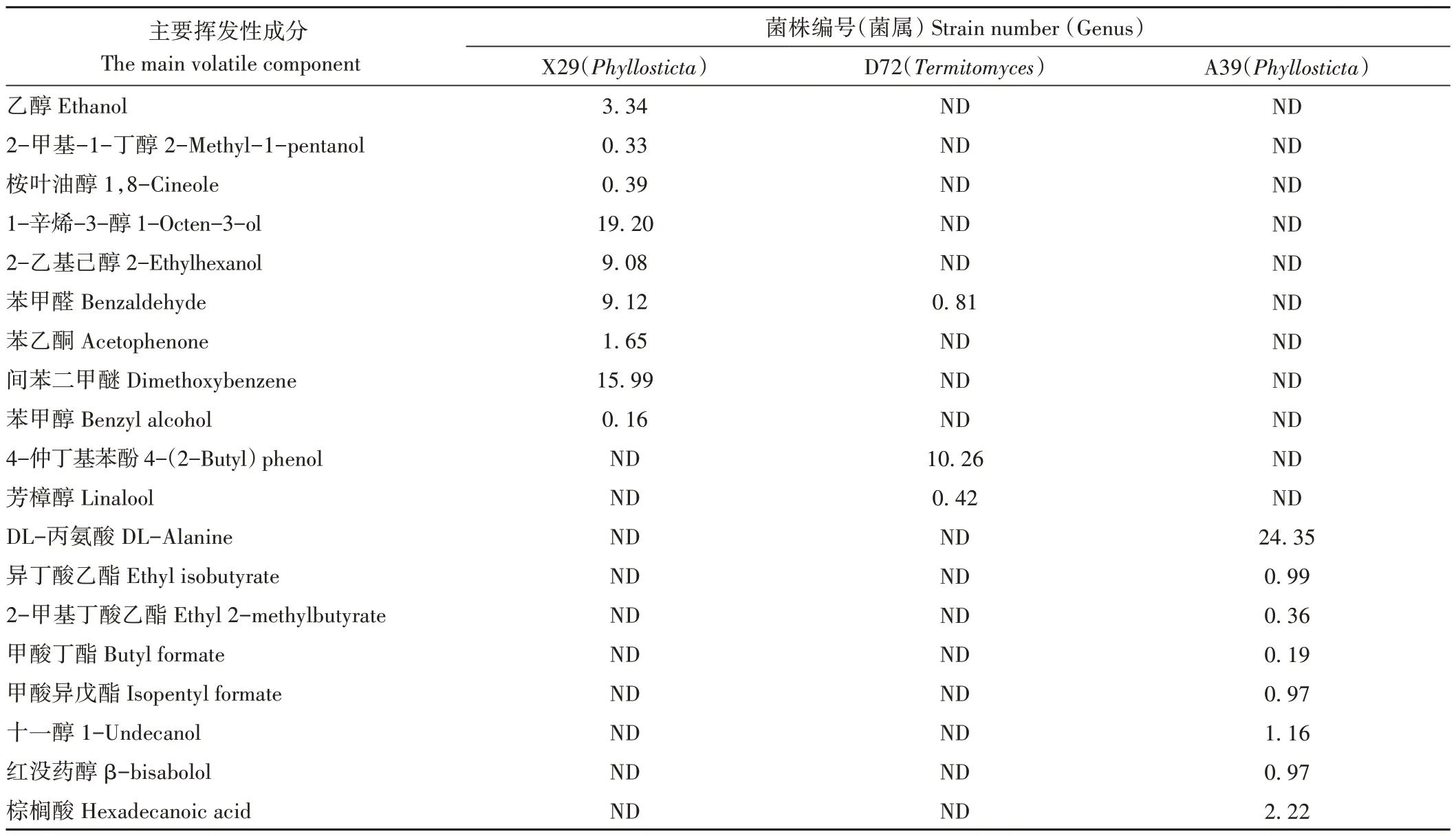
表5 3株产精油成分菌株的主要挥发性物质含量Table 5 Contents of main volatile components in three strains producing essential oil/%
3 讨论
内生真菌是参与宿主植物生长、发育及代谢等过程的重要因子,为开发潜在菌株并分析与宿主功能的相关性,采用传统分离培养法对不同株系和不同季节油樟叶内生真菌进行分离纯化和鉴定,可知菌株数量和种属随株系和季节的不同而不同,其中16 号株系和春季菌群多样性相对最丰富,53号和冬季次之,1 号和秋季最低。周宁等[27]发现不同季节蔗叶内生真菌多样性和分布存在较大差异。吕立新等[28]分析春、夏和秋茅苍术叶的内生真菌群落分布,表明菌群随季节的变化具有一定的演替规律,夏季真菌多样性最高。周万海等[5]研究了不同季节油樟叶内生细菌的群落组成,发现秋季多样性种群较丰富且分布均匀,冬季菌株丰富度和均匀度均较低。因此,菌株多样性可能因宿主生长环境、品种、株龄等不同而不同,且内生真菌和内生细菌具有非一致的变化趋势。由蔡媛等[29]报道可知多花黄精中分离的内生细菌鉴定为19 门419 属,而其内生真菌仅鉴定为7 门187 属。任婷月等[30]采用高通量测序技术对3 个不同品种大麦内生真菌和细菌进行测定,显示不同品种的内生菌群落组成差异均较大,与本试验结果相似。内生真菌群落结构与植株状态、生长和代谢关联。杨琴等[31]研究显示不同树龄银杏叶的优势菌门均为子囊菌门,曲霉属、假丝酵母属、球腔菌属等为优势菌属。Wu 等[32]研究发现艾蒿叶片内生真菌优势属为担孢酵母属(Erythrobasidium)和掷孢酵母属(Sporobolomyces)。本试验分离的43 株内生真菌优势门为Ascomycota,优势属为Phyllosticta,与前人研究具有较大差异。同时,本实验室前期利用高通量测序技术对秋季油樟叶片内生真菌类群进行分析,显示优势属为子囊菌门未分类属(unclassified Ascomycota)、钉孢属(Passalora)和马氏霉属(Uwebraunia)[33];而本研究中来自秋季油樟叶的菌株仅有2 株,隶属于Alternaria和Phyllosticta,推测是由于受到培养方法、培养基类型和条件局限性所致。
部分内生菌能够增强宿主植物的健康状态并促进分泌有效的代谢产物[34],通过与宿主共生诱导能产生与宿主植物相同或相似的代谢产物,如紫锥菊[35]茎叶组织分离的内生细菌可以影响植物挥发性产物和生物碱的合成。暗隔内生菌能促进水稻生长、营养物质的吸收并增强抗逆性[36]。本研究中,虽然春季16号株系油樟叶产油量最高,且内生真菌丰富度、均匀度最佳,但Pearson 相关分析发现,不同株系或不同季节油樟叶精油产率与其中可培养内生真菌多样性指数无相关性,这与严宽等[21-22]的研究结果不一致;上述结果的差异可能与研究材料形态特征不同有关,亦可能与宿主不同株系或环境下含有的特定内生菌诱导产生特定代谢物有关,后续应对不同株系或不同季节油樟叶精油主要成分与内生真菌多样性进行分析,以确定挥发油含量和成分如何影响其中内生真菌。本研究进一步分析还发现,Penicillium和Cladosporium为16号株系特有属,Cladosporium、Phomopsis、Diaporthe为春季特有属,属于此分类的菌株可能能促进油樟叶精油中特定成分的合成进而提高精油的积累,但相关机制后续还应深入探究。
目前关于菌株的挥发性成分研究主要集中于抗菌性、香气和病虫害防治等方面,如杨黎华等[37]从香茅草分离的内生真菌中筛选得到1 株长喙壳属菌株,其发酵液具有清甜香和花香,利用气相色谱/质谱仪鉴定出乙缩醛二乙醇、柠檬烯、月桂烯、月桂醛等74种香料物质;冯福山等[38]发现枯草芽胞杆菌Y13 发酵液产生的挥发性物质对油茶炭疽病菌具有较好生防效果;陶玲芸等[39]发现棘孢木霉挥发性次级代谢产物中6-正戊基-2H-吡喃-2-酮(6-PAP)成分能抑制尖孢镰孢菌。本试验从油樟叶中提取的精油具有特殊芳香气味,通过GC/MS对内生真菌发酵液进行分析鉴定,可知菌株能产生具有醇类、酸类、酯类、烯类、酮类、醛类及酚类等69 种主要挥发性成分。其中苯乙酸乙酯具有浓烈而甜的蜂蜜香气;甲酸乙酯具有辛辣的刺激味和菠萝样的果香香气;6-戊基-2H-吡喃-2-酮可用于食品用香料,呈蘑菇、蓝干酪或乳品香气;对甲基苯酚呈烟熏、草药气味,尤其是桉叶油醇、芳樟醇和红没药醇,它们属于油樟精油成分,说明油樟内生真菌产生与宿主相同的产物,如X29、D72、A39 菌株,且X29 和A39 均为Phyllosticta,但含量较少,后续将展开进一步条件优化提高产量,并探索其功能。
4 结论
本研究采用传统微生物纯培养法研究了四川宜宾地区特色香料作物油樟叶部内生真菌群落的结构和多样性,并测定其产油量和菌株发酵液挥发性成分。结果表明,不同株系和不同季节油樟叶的内生真菌在数量、种群及组成上存在差异,油樟叶中可培养内生真菌是一个重要的资源库,其中16 号株系和春季油樟叶内生真菌群落的多样性相对最丰富。通过产油率和可培养内生真菌多样性指数关联分析,证实两者无显著相关关系。此外通过菌株发酵液成分分析,鉴定到醇类、酯类、烯类及酮类等化合物,并显示菌株能分泌产生精油类代谢产物,具有进一步开发香料成分的潜力。后续应加大对微生物功能的探索,对功能性菌株开展应用研究。
