苦参总黄酮对D-半乳糖致神经细胞衰老的改善作用及相关机制
2024-02-22马亚飞
马亚飞
(河南省中医院药学部,河南 郑州 450000)
阿尔茨海默病、帕金森病等神经退行性疾病均表现出年龄依赖性〔1〕,神经细胞衰老是神经退行性病变的主要诱因,促进衰老的因子可能参与神经退行性疾病的发生与发展,延缓衰老有助于降低发病风险〔2〕。衰老的机制十分复杂,包括表观遗传改变、基因稳定性降低、端粒缩短、线粒体功能障碍、蛋白水解遗传等,氧化应激在其中发挥重要作用〔3〕。近年来研究发现,总黄酮可通过调节肝癌细胞中超氧化物歧化酶、过氧化氢酶等氧化应激关键因子表达,降低活性氧水平,进而发挥抗氧化作用〔4〕;此外总黄酮还可通过清除氧自由基、抑制脂质过氧化来延缓机体衰老过程〔5〕。生物碱和黄酮是苦参的主要有效成分,目前已对生物碱类成分进行了较多研究报道,评价苦参的质量也主要通过生物碱含量来决定〔6〕,但对于黄酮类成分的研究仍十分少见。本研究旨在探讨苦参总黄酮对D-半乳糖致神经细胞衰老的改善作用及相关机制。
1 资料与方法
1.1细胞与主要试剂 PC12神经元细胞购于北京伊莱瑞生物科技有限公司;苦参总黄酮(批号:ST15692)购于成都彼样生物科技有限公司。DMEM培养基、胎牛血清购于哈灵(上海)生物科技有限公司;D-半乳糖购于美国Sigma公司;超氧化物歧化酶、丙二醛、乳酸脱氢酶检测试剂盒购于上海舜冉生物科技有限公司;鼠抗人B淋巴细胞瘤(Bcl)-2相关X蛋白(Bax)单克隆抗体、兔抗人细胞色素(Cyt)-C单克隆抗体、鼠抗人Bcl-2单克隆抗体、鼠抗人核因子E2相关因子(Nrf)2单克隆抗体、兔抗人血红素氧合酶1单克隆抗体、鼠抗人GAPDH单克隆抗体及羊抗鼠IgG、羊抗兔IgG购于天津卡梅德生物科技有限公司。
1.2PC12神经元细胞衰老模型建立与分组 PC12神经元细胞用DMEM完全培养基常规培养,取传至第4代的细胞按2×108个/L,每孔200 μl的密度接种到96孔板中,用含D-半乳糖的完全培养基孵育24 h后更换为完全培养基继续孵育,D-半乳糖终浓度为40 mg/ml。造模成功后分为模型组,10、20、40 μg/ml苦参总黄酮组,每组10个复孔,同时设置空白对照组。空白对照组使用正常PC12细胞在完全培养基中培养;模型组使用完全培养基培养;不同剂量苦参总黄酮组完全培养基中分别加入终浓度为10、20、40 μg/ml的苦参总黄酮。上述各组继续培养48 h后进行后续实验。
1.3细胞存活率、衰老细胞情况、凋亡率检测 四甲基唑蓝(MTT)法检测各组细胞存活率,细胞存活率=(苦参总黄酮组或模型组吸光度值-阴性对照组吸光度值)/(空白对照组吸光度值-阴性对照组吸光度值)×100%。β-半乳糖苷酶染色试剂盒检测各组细胞衰老情况,光学显微镜下计数染蓝色的衰老细胞数目,计算β-半乳糖苷酶阳性染色率。细胞凋亡检测:向各组细胞中加入膜联蛋白Ⅴ(Annexin Ⅴ)结合缓冲液(10倍)工作液将细胞重悬,加入Annexin V-PE/7-AAD荧光双染色剂各5 μl,避光静置20 min,流式细胞仪检测细胞凋亡率。
1.4细胞周期分布情况检测 取各组细胞接种到96孔板中,孵育24 h,2 000 r/min离心5 min,弃去上清,磷酸盐缓冲液重悬细胞,再次离心后弃去上清,轻弹离心管底部将细胞分散,避免成团聚集,加入预冷的70%酒精1 ml,吹打混合均匀,4 ℃固定40 min,2 000 r/min离心5 min,弃去上清。向离心管中加入碘化丙啶(PI)染色液500 μl重悬细胞沉淀,37 ℃避光静置30 min,使用流式细胞仪检测细胞周期。
1.5各组细胞抗氧化指标检测 取各组细胞制成约10%匀浆液,放于-20 ℃中冰融3次,光学显微镜下观察细胞破碎情况,若破碎不完全继续冰融2次;3 000 r/min离心10 min,收集上清液,按照试剂盒检测说明书测定超氧化物歧化酶、丙二醛、乳酸脱氢酶浓度。
1.6Western印迹检测各组细胞中Bax、Cyt-C、Bcl-2、Nrf2、血红素氧合酶1蛋白表达水平 RIPA细胞裂解液提取各组细胞总蛋白,二喹啉甲酸(BCA)试剂盒测定蛋白浓度,电泳后将目标蛋白转移至聚偏氟乙烯(PVDF)膜,封闭2 h,加入鼠抗人Bax单克隆抗体(1∶1 000稀释)、兔抗人Cyt-C单克隆抗体(1∶1 000稀释)、鼠抗人Bcl-2单克隆抗体(1∶500稀释)、鼠抗人Nrf2单克隆抗体(1∶500稀释)、兔抗人血红素氧合酶1单克隆抗体(1∶1 000稀释)、鼠抗人GAPDH单克隆抗体(1∶2 000稀释),4 ℃孵培育过夜,加入二抗:辣根过氧化物没标记的羊抗鼠IgG、羊抗兔IgG,均为1∶4 000稀释,室温下孵育1 h,电化学发光(ECL)显影,使用ImageJ软件读取各条带的灰度值。
1.7统计学分析 采用SPSS25.0软件进行方差分析和t检验。
2 结 果
2.1各组细胞存活率、衰老细胞情况、凋亡率比较 模型组细胞存活率明显低于空白对照组,β-半乳糖苷酶阳性染色率、细胞凋亡率明显高于空白对照组(P<0.01);苦参总黄酮组细胞存活率明显高于模型组,β-半乳糖苷酶阳性染色率、细胞凋亡率明显低于模型组,且随着苦参总黄酮给药浓度的增加变化逐渐明显,差异有统计学意义(P<0.05)。见表1。
2.2各组细胞周期分布情况比较 各组G0/G1期、S期细胞比例差异有统计学意义(P<0.01);模型组G0/G1期细胞比例明显高于空白对照组,S期细胞比例明显低于空白对照组(P<0.01);苦参总黄酮组G0/G1期细胞比例明显低于模型组,S期细胞比例明显高于模型组(P<0.05)。见表1。

表1 各组细胞存活率、衰老细胞、凋亡率及细胞周期分布比较
2.3各组细胞抗氧化指标水平比较 各组超氧化物歧化酶、丙二醛、乳酸脱氢酶浓度差异有统计学意义(P<0.05,P<0.001);模型组超氧化物歧化酶浓度明显低于空白对照组,丙二醛、乳酸脱氢酶浓度明显高于空白对照组(P<0.01);苦参总黄酮组超氧化物歧化酶浓度明显高于模型组,丙二醛、乳酸脱氢酶浓度明显低于模型组,且随着苦参总黄酮给药浓度的增加变化逐渐明显,差异有统计学意义(P<0.05)。见表2。
2.4各组细胞中Bax、Cyt-C、Bcl-2蛋白表达水平比较 各组Bax、Cyt-C、Bcl-2蛋白表达水平差异有统计学意义(P<0.01);模型组Bax蛋白表达水平明显高于空白对照组,Cyt-C、Bcl-2蛋白表达水平明显低于空白对照组,差异有统计学意义(P<0.01);苦参总黄酮组Bax蛋白表达水平明显低于模型组,Cyt-C、Bcl-2蛋白表达水平明显高于模型组,且随着苦参总黄酮给药浓度的增加变化逐渐明显,差异有统计学意义(P<0.05)。见表2、图1。
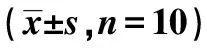
表2 各组细胞抗氧化指标水平及细胞中Bax、Cyt-c、Bcl-2蛋白表达水平比较
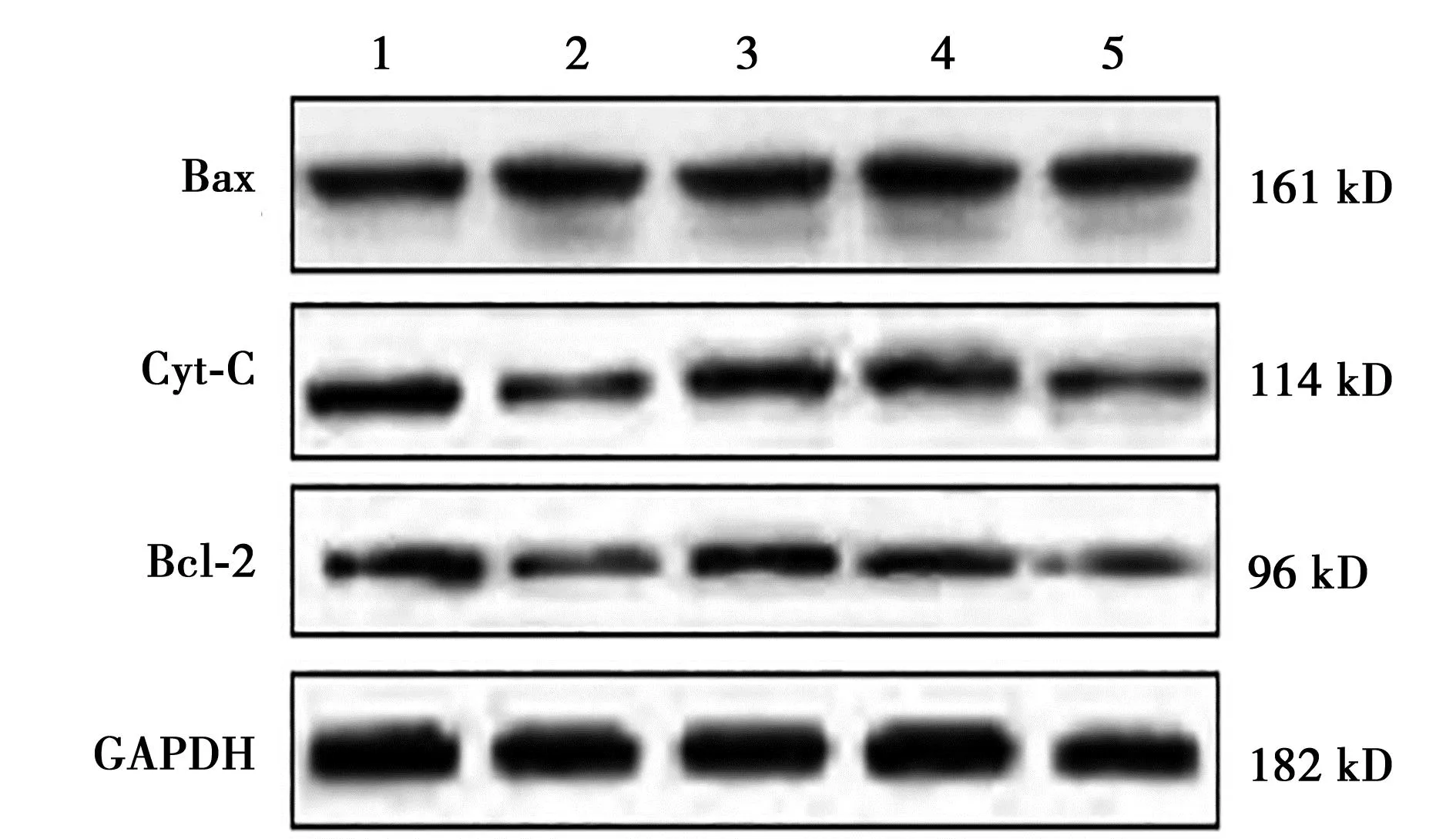
1~5:空白对照组、模型组、10、20、40 μg/ml苦参总黄酮组;下图同图1 各组细胞中Bax、Cyt-C、Bcl-2蛋白表达
2.5各组细胞中Nrf2、血红素氧合酶1蛋白表达水平比较 各组Nrf2、血红素氧合酶1蛋白表达水平差异有统计学意义(P<0.01);模型组Nrf2、血红素氧合酶1蛋白表达水平明显高于空白对照组(P<0.01);苦参总黄酮组明显低于模型组,且随着苦参总黄酮给药浓度的增加变化逐渐明显,差异有统计学意义(P<0.05)。见图2、表3。

图2 各组细胞中Nrf2、血红素氧合酶1蛋白表达
表3 各组细胞中Nrf2、血红素氧合酶1蛋白表达水平比较
3 讨 论
正常生理状态下D-半乳糖经肝脏代谢,当其浓度异常升高时会逐渐积累,造成细胞渗透压改变、肿胀及细胞膜脂受损,还能够与胺相互作用,生成高级糖基化终产物,引发细胞衰老〔7,8〕。本研究使用D-半乳糖建立PC12神经元细胞衰老模型。氧化应激是引发细胞衰老的主要原因之一,主要由抗氧化反应系统与自由基生成水平之间平衡失调所致,中枢神经系统对氧化应激损伤的敏感度最高。近年来研究发现,总黄酮具有抗氧化作用,能够防止细胞退化、衰老;其中金雀异黄酮对衰老H9c2细胞具有保护作用〔9〕;欧亚旋覆花总黄酮能够对抗L929细胞衰老和自噬〔10〕;但关于苦参总黄酮对神经元细胞衰老影响的研究仍十分少见。
本研究结果提示,苦参总黄酮能够提高衰老神经细胞的存活率,减少衰老神经细胞数量,降低细胞凋亡率,且呈剂量依赖性,随着给药浓度的增加作用效果更加明显。细胞衰老时细胞周期会停留在G1期,细胞快速增殖时细胞周期会停留在S期〔11〕。本研究结果说明苦参总黄酮能够有效降低停留在G1期的细胞比例,升高S期细胞比例,进而提高衰老细胞的增殖活力。抗氧化应激能力直接影响到对抗细胞衰老的效果,超氧化物歧化酶参与维持机体的氧化与抗氧化平衡〔12〕。当机体存在大量超氧自由基时会与膜脂质反应生成丙二醛,过量累积的丙二醛会损伤细胞膜的正常结构和功能〔13〕。乳酸脱氢酶是一种氧化还原酶,与氧化应激水平密切相关〔14〕。结合本研究结果可知苦参总黄酮可通过升高超氧化物歧化酶水平,降低丙二醛、乳酸脱氢酶水平来降低衰老神经细胞的氧化损伤程度。
细胞凋亡是一个多种因子共同参与的复杂过程,其中促凋亡因子Bax、Cyt-C与抗凋亡因子Bcl-2平衡失调发挥重要作用〔15〕。本研究结果提示,苦参总黄酮可通过下调Bax蛋白表达,上调Cyt-C、Bcl-2蛋白表达来降低衰老神经细胞的凋亡率。Nrf2是细胞氧化应激反应的转录因子,具有很高的敏感性,同时也是抗氧化防御细胞的调控因子,与下游抗氧化酶表达水平密切相关〔16〕。血红素氧合酶1是Nrf2信号通路下游的抗氧化蛋白,能够降低氧化应激损伤,抑制神经细胞凋亡〔17〕。本研究提示,苦参总黄酮可通过促进Nrf2表达来提高下游抗氧化蛋白血红素氧合酶1水平,进而抵抗氧化应激的发生,改善D-半乳糖诱导的神经细胞衰老,在神经退行性疾病防治中具有潜在的应用价值。
