牛环形泰勒虫烯醇化酶重组蛋白对小鼠免疫应答的研究
2024-01-29郑会珍何文文葛晓敏李永畅巴音查汗郭庆勇
郑会珍,甘 露,刘 燕,何文文,葛晓敏,李永畅,巴音查汗,郭庆勇
(新疆农业大学动物医学学院,新疆乌鲁木齐 830052)
牛环形泰勒虫病急性感染时可致宿主高热、贫血、黄疸及体表淋巴结肿大等症状[1-2]。高发病率和高病死率导致产奶量及产肉量降低,给养牛业带来严重的经济损失[3]。目前,环形泰勒虫病的防治主要通过疫苗预防、药物除蜱和药物治疗3个方面,但疫苗预防和药物除蜱存在不能提供交叉保护效果、蜱虫不易完全除去且存在药物残留等弊端[4-7]。目前,针对泰勒虫病最有效的药物是羟基萘醌家族(帕伐醌和布帕伐醌),但国内外均出现对布帕伐醌耐药虫株的报道[8]。综上所述,布帕伐醌在未来很可能无法进行有效使用,人们急需研制具有不同作用机制的新型药物或化合物。烯醇化酶(enolase)在糖酵解中通过催化2-磷酸甘油酸形成磷酸烯醇式丙酮酸为宿主提供能量,且在多种生物体内高度保守[9]。而且,enolase作为药物靶点和候选疫苗的可能性已经在疟原虫、锥虫、泰勒虫中被发现[10-11],对其保护性抗原特性的深入研究可使人们更清楚其在寄生虫入侵宿主、供能中的相关作用,也为潜在的疫苗候选抗原、诊治方法和药物筛选研究奠定基础。本试验将牛环形泰勒虫烯醇化酶重组蛋白(rTa-enolase)诱导表达,纯化后免疫Balb/c小鼠,用ELISA、MTT和流式细胞术检测免疫动物特异性抗体水平和细胞亚型比例等指标,评估rTa-enolase对试验动物体液免疫水平和细胞免疫水平的影响,为后续牛环形泰勒虫保护性候选抗原、亚单位疫苗及药物靶点的相关研究提供基础。
1 材料与方法
1.1 材料
1.1.1 血清和实验动物 环形泰勒虫标准阳性和阴性血清,由新疆农业大学动医学学院寄生虫实验室提供;清洁级6周龄Balb/c小鼠,购自新疆医科大学实验动物中心。
1.1.2 菌株 pET-32a-enolase菌株由新疆农业大学动医学学院寄生虫实验室提供。
1.1.3 试剂 His-tag Purification Resin,上海碧云天生物技术有限公司产品;弗氏完全佐剂、弗氏不完全佐剂以及质粒小剂量提取试剂盒、RPMI1640 ,北京鼎国昌盛生物技术有限责任公司产品;ConA,Sigma公司产品;MTT 细胞增殖及细胞毒性检测试剂盒、PE Anti-Mouse CD3 Antibody、FITC Anti-Mouse CD8 Antibody、PE/Cyanine5.5 Anti-Mouse CD4 Antibody,武汉伊莱瑞特生物科技股份有限公司产品;Anti-Mouse CD16/CD32 (2.4G2),Purified流式抗体、Flow cytometry Staining buffer流式染色缓冲液,杭州联科生物技术股份有限公司产品。
1.1.4 主要仪器 超净工作台(SW-SJ-2D),苏州净化设备有限公司产品台式;恒温振荡器,苏州培英实验设备有限公司;超声细胞粉碎机(JY92-IIN),宁波新芝生物科技有限公司产品;稳压电泳仪(DYY-Ⅲ-5),北京六一生物科技有限公司产品;酶标仪(ST-360),上海科华生物工程有限公司产品;CO2生化培养箱,美国Eppendorf公司产品;流式细胞仪(S3e),Bio-Rad公司产品。
1.2 方法
1.2.1 牛环形泰勒虫烯醇化酶原核重组蛋白的表达、纯化及鉴定 将原核阳性菌液pET-32a-enolase复苏后,在200 mL LB培养基(Amp+)扩大培养,待菌液OD 600 nm值达到0.4~0.6时,加入0.6 mmol/L IPTG在25 ℃诱导5 h,收集菌液沉淀,超声破碎,利用His-tag进行蛋白纯化、超滤管复性浓缩,使用单滴分度计测得蛋白浓度通过SDS-PAGE鉴定后以环形泰勒虫阳性血清(1∶100)作为一抗、兔抗牛 IgG-HRP(1∶10 000)作为二抗,经Western blot鉴定重组蛋白的表达。
1.2.2 免疫小鼠IgG/IgG1/IgG2a抗体水平检测 利用方法1.2.1纯化重组蛋白rTa-enolase免疫Balb/c小鼠,免疫前尾尖采血后分离血清,作为阴性对照。将纯化后重组蛋白与弗式佐剂等体积混合,超声破碎后成油包水状,皮下多点注射。每隔2周使用弗式不完全佐剂进行第2次免疫和第3次免疫,免疫得蛋白量分别为每只小鼠一免200 μg、二免和三免100 μg免疫。3次免疫后10 d,经心脏采血并分离血清。采用间接ELISA法测定抗体水平,将纯化后的重组蛋白(2 μg/L)过夜包被96孔酶标板,以制备的鼠多克隆抗体倍比稀释(1∶200~1∶6 553 600)为一抗,以山羊抗鼠HRP-IgG/IgG1/IgG2a(1∶10 000)为二抗,使用酶标仪检测OD 450 nm值。
1.2.3 MTT检测小鼠脾脏淋巴细胞再刺激后细胞增殖情况 随机选取3只小鼠眼眶摘眼球处死,无菌取脾脏组织,制备脾细胞,调整细胞浓度为每孔浓度1×106个/mL每孔100 μL 细胞悬液铺于96孔板中。加入3组刺激剂:每孔20 μL的PBS、每孔0.005 g/L的rTa-enolase重组蛋白、以及0.01 g/L的ConA。置于37 ℃体积分数为5%的CO2生化培养箱中72 h。每孔再加50 μL 1×MTT工作液,于37 ℃培养箱中孵育4 h,小心吸出上清液,每孔加150 μL的DMSO使甲臜溶解,用平板摇床摇匀。使用酶标仪检测OD 570 nm值。检测结果以刺激指数(SI)表示SI=OD实验孔值/OD对照孔值
1.2.4 流式细胞术检测小鼠脾脏混合淋巴细胞亚群 用200 μL的细胞染色缓冲液重悬1×106个细胞,每管加入2 μL的anti-mouse CD16/32 抗体于4 ℃冰箱中封闭10 min;避光加入PE Anti-Mouse CD3 Antibody、FITC Anti-Mouse CD8 Antibody、PE/Cyanine5.5 Anti-Mouse CD4 Antibody 4 ℃孵育40min,离心,弃上清。加入预冷的细胞染色缓冲液洗2遍,1 800 r/min 4 ℃ 离心 5 min后弃掉上清,加入细胞染色缓冲液将细胞悬起并转移到预先做好标记的流式管中使用流式细胞仪检测CD4+T/CD8+T细胞比值。
1.2.5 数据处理 数据利用Spss单因素ANOVA检验进行差异显著性分析,P<0.05为差异具有显著统计学意义,P<0.01为差异具有极显著统计学意义,借助Flow Jo、GraphPad Prism 9.1.0等软件对所得数据进行分析和制图。
2 结果
2.1 牛环形泰勒虫烯醇化酶原核重组蛋白的表达、纯化及鉴定
重组蛋白纯化后经SDS-PAGE凝胶电泳验证后获得大小为68 ku的单一目的蛋白(图1A)。Western blot结果显示,Enolase可与牛环形泰勒虫阳性血清发生反应,在68 ku处出现特异性条带,阴性血清不发生反应(图1B)。
2.2 小鼠血清中IgG抗体检测结果
纯化后的rTa-enolase为免疫原,皮下多点注射Balb/c小鼠免疫3次(每隔14 d免疫)后采血,采用间接ELISA法测定免疫血清中抗体水平,结果显示一免后14 d抗体水平达到1∶51 200,完全免疫后小鼠的特异性抗体水平高达1∶6 553 600(图2A),且免疫前后抗体水平差异极显著(P<0.01)。
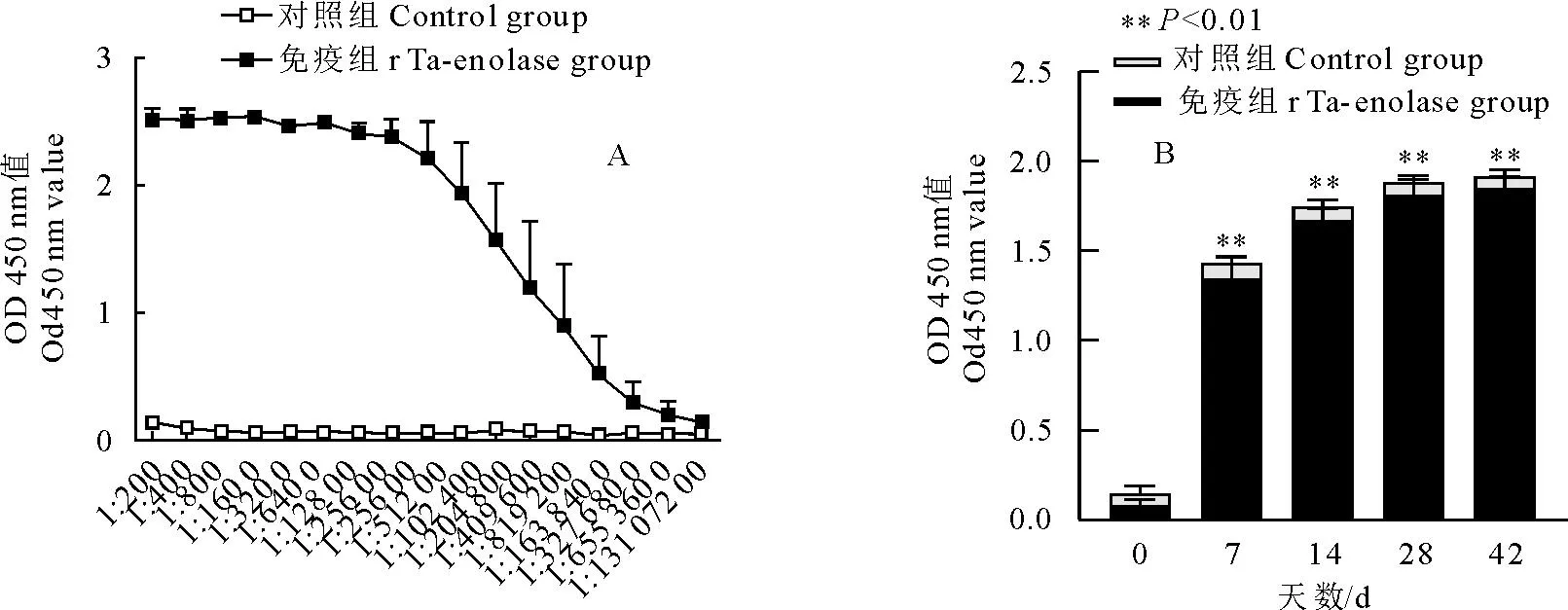
A.三次免疫后小鼠抗体水平; B.免疫前后小鼠IgG抗体水平A.Antibody levels of mice after three times of immunization; B.Antibody levels in mice before and after immunization图2 rTa-enolase免疫小鼠后血清抗体水平检测Fig.2 Serum antibody levels of mice immunized with rTa-enolase
2.3 小鼠血清中IgG1/IgG2a抗体检测结果
采用间接ELISA测定免疫血清中抗体亚型IgG1和IgG2a水平,结果显示免疫后小鼠IgG1和IgG2a均与对照组差异极显著(P<0.01),免疫后IgG1极显著高于IgG2a水平(P<0.01)(图3)。

A.IgG1; B.IgG2a; C.免疫前后小鼠IgG1与IgG2a抗体水平差异性分析A.IgG1;B.IgG2a;C.Analysis of the difference of IgG1 and IgG2a antibody levels in mice before and after immunization图3 rTa-enolase免疫小鼠后血清特异性抗体IgG1/IgG2a检测Fig.3 Detection of serum antibody subtype IgG1/IgG2a in mice immunized with rTa-enolase
2.4 MTT法检测小鼠脾脏混合淋巴细胞在刺激后细胞增殖
使用MTT细胞增殖及细胞毒性检测试剂盒检测体外刺激小鼠后的细胞增殖情况,结果如图4显示Con A组增殖指数为3.96±0.68、大于2.0代表说明整个实验系统可靠,rTa-enolase刺激组为2.61±0.86,与对照组(1.07±0.03)对比差异显著(P<0.05),可明显促进小鼠淋巴细胞增殖(图4)。
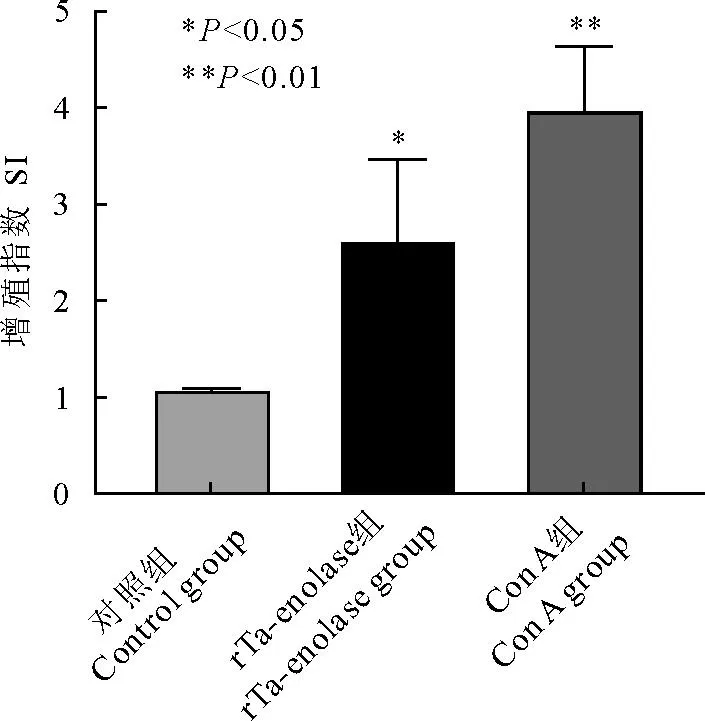
图4 MTT法检测脾脏淋巴细胞体外刺激后细胞增殖情况Fig.4 Detection of the proliferation of splenic lymphocytes after in vitro stimulation by MTT assay
2.5 流式细胞术检测小鼠脾脏混合淋巴细胞
3次免疫后,免疫组和对照组小鼠脾脏淋巴细胞CD4+/CD8+T细胞的比值分别为2.53±0.11和1.85±0.11,差异极显著(P<0.01),单独的淋巴细胞亚群比例比较显示,CD4+T细胞rTa-enolase免疫组比值(52.70±1.08)极显著高于对照组(47.33±1.46)(P<0.01),CD8+T淋巴细胞免疫组显著低于对照组(P<0.01)(图5)。

A.对照组; B.rTa-enolase免疫组; C.脾脏中T淋巴细胞分析A.Control group; B.rTa-enolase immune group; C.Analysis of T lymphocytes in spleen图5 免疫小鼠淋巴细胞亚群分布Fig.5 Distribution of lymphocyte subsets in immunized mice
3 讨论
环形泰勒虫病的高病死率、治疗和控制措施的高成本等负面影响给养牛业造成了巨大的经济损失。目前,耐药性又成为环形泰勒虫预防和控制的难题之一,人们迫切需要寻找预防和控制病原体新型方法。研究表明烯醇化酶在环形泰勒虫葡萄糖代谢中起着至关重要的作用,而且近年的研究发现,enolase的抑制剂(香豆素衍生物)可显著抑制牛环形泰勒虫而不抑制宿主,可作为一种潜在的药物蛋白靶点[12]。
IgG抗体是血清中含量最多的抗体,且持续时间长,是机体抗感染免疫的主力,也是介导体液免疫的主要抗体[13]。本试验免疫小鼠IgG抗体含量始终高于对照组,表明rTa-enolase在免疫小鼠中能诱导出良好、持久的抗体水平,说明该蛋白可以引起良好的体液免疫反应。与前人研究[14]相比,本试验重组蛋白rTa-enolase免疫小鼠后获得相当高水平的体液免疫效果。
Th2细胞诱导特异性IgG1型抗体增加,介导体液免疫,Th1细胞通过分泌细胞因子诱导IgG2a抗体的产生,增强免疫反应的识别阶段[15]。研究表明,清除感染小鼠体内的锥虫[16]和弓形虫[17]需要Th1型免疫反应。本研究发现,免疫后IgG抗体亚型IgG1和IgG2a均与对照组差异极显著(P<0.01),而且免疫后IgG1极显著高于IgG2a水平(P<0.01),说明本试验重组蛋白可引起Th1与Th2型联合免疫反应,免疫IgG1引起的Th2型反应占据主导地位,结合免疫后产生极高的IgG抗体水平,由此推测,enolase可能可以通过Th1型结合Th2型免疫反应来保证机体免受病原的入侵。体外培养时,T细胞可通过特定抗原或促丝分裂原剂的刺激,将小淋巴细胞转化为淋巴细胞,T淋巴细胞的转化率反映了体内的细胞免疫水平[18]。本研究免疫组的T淋巴细胞增殖指数显著高于对照组(P<0.05),与Xue S等[15]研究支原体enolase在实验动物中诱导的免疫反应一致,均可明显诱导细胞增殖,刺激机体T细胞转化为淋巴细胞,从而产生较好的细胞免疫反应。
另外,T淋巴可通过CD4+T淋巴细胞(Th细胞)和CD8+T淋巴细胞(Tc细胞)亚群释放大量细胞因子增强机体免疫应答水平,CD4+T淋巴细胞的增多可促进机体分泌Th1型细胞因子,激活巨噬细胞、NK细胞和消除寄生虫[19],增强机体细胞免疫应答功能。本研究利用流式细胞术将免疫细胞通过其表面荧光进行T细胞亚型分析,结果表明,本试验免疫后小鼠的CD4+T细胞、CD4+T/CD8+T细胞比值与对照组相比差异显著(P<0.01),说明该重组蛋白能促进CD4+淋巴细胞的增多来促进细胞因子的分泌,来达到增强小鼠细胞免疫功能。Zhang X等[20]在旋毛虫真核重组和原核重组enolase蛋白免疫小鼠也诱导类似的免疫反应。
综上所述,rTa-enolase重组蛋白的CD4+T细胞的分化情况可能为机体可诱导机体体液和细胞免疫应答,未造成机体的免疫抑制或过度反应,可为宿主提供免疫保护作用,为牛环形泰勒虫候选疫苗、药物靶点的临床试验等相关研究提供了数据支持。
