miR-494靶向TLR-4通路在骨质疏松发病中的作用及机制*
2024-01-10匡嘉兵雷建平程煜方
匡嘉兵 郭 松 雷建平 程煜方
(武汉市第四医院骨科,武汉 430060)
社会和经济的发展带来人口老龄化的增加以及生活方式的改变,进而导致骨质疏松症骨代谢紊乱的发病率逐年增加[1]。根据世界卫生组织的定义,骨质疏松症的特征主要包括骨量的丢失和骨微结构受到破坏。骨质疏松性骨折往往会给老年人造成长期行动不便和各种并发症,影响患者的生活质量甚至生命健康[2]。因此骨质疏松症逐渐成为世界范围内的研究热点,但目前对于骨质疏松的发病机制尚未完全明确。
微小RNA(microRNA)是单链非编码RNA,包含20~22个核苷酸。microRNA通过靶向靶基因的3′UTR片段并提示其表达,被认为是多种细胞过程的潜在调节因子,如细胞增殖、凋亡和细胞分化[3]。最近的研究概述了数千个microRNA和参与调控它们的多种人类基因。许多研究证实了各种microRNA在调节骨质疏松中的作用[4-8]。先前有研究表明,miR-30a-5p可通过调控RUNX2的表达,抑制成骨细胞分化[6]。miR-204通过调控Wnt信号通路影响小鼠成骨细胞分化等[7]。我们先前的研究证实,miR-494抑制骨钙素活性,抑制成骨细胞形成矿化结节,但miR-494对骨质疏松的影响尚未有体内实验进行证实和研究[8]。Toll样受体-4(TLR4)通路与影响成骨细胞的其他重要通路存在密切联系。我们经过先前的研究已经证实miR-494可能调控TLR4的表达,进而抑制骨钙素活性,抑制成骨细胞形成矿化结节[8]。
我们推测miR-494可能在骨质疏松发病过程中起作用,并可能通过靶向TLR-4参与骨质疏松大鼠骨髓间质细胞的增殖、成骨分化、成脂分化等多种重要过程。为深入探究miR-494在骨质疏松中的发病机制,通过构建骨质疏松大鼠模型进行体内实验,探究miR-494对TLR4通路的影响以及在骨质疏松发病中的作用及机制。
1 材料和方法
1.1 材料
选取30只SPF级8周龄雌性Sprague-Dawley大鼠,购自南方医科大学。生产许可证【SCXK(粤)2021-0041】,使用许可证【SYXK(鄂)2021-0114】,大鼠平均体质量为(218.74±23.18)g。
1.2 方法
1.2.1实验分组:将入组的大鼠随机分为NC组(阴性对照大鼠)、OP组(骨质疏松大鼠)、OP+miR-494 inhibitor组(骨质疏松大鼠,miR-494抑制物注射)、OP+miR-494 inhibitor+TLR-4 vector组(骨质疏松大鼠,miR-494抑制物和TLR-4过表达腺病毒载体注射)、OP+miR-494 inhibitor+sh TLR-4组(骨质疏松大鼠,miR-494抑制物和TLR-4沉默腺病毒载体注射),每组6只。本研究获得本院动物伦理委员会批准,伦理审批号:LLX20210301D。
1.2.2模型构建:入组的NC组大鼠以每天70 mg/kg的0.9%氯化钠溶液进行灌胃,每两天称量一次体质量,连续灌胃两周。其余组大鼠均构建骨质疏松大鼠模型,以每天70 mg/kg维甲酸溶液进行灌胃,连续灌胃两周即可造模成功。麻醉后,首先从所有大鼠的腹主动脉抽取血液。每组6只大鼠通过多聚甲醛灌注固定,获得股骨组织。
1.2.3骨标本及血液样本采集及制备:各组大鼠腹腔注射3%戊巴比妥钠按照0.17 mL/100 g的标准麻醉,采集各组大鼠腹主动脉血,将腹主动脉血在3000 r/min的高速离心机中离心10 min,并获得上清液,置于-4 ℃冰箱保存。随后,对大鼠四肢皮肤进行初步消毒和切断,采用断颈法处死所有大鼠,取双侧股骨,剔除肌肉和筋膜,用生理盐水纱布,锡纸包裹,左侧用于分离BMSCs细胞,右侧用于测量骨密度。
1.2.4分离BMSCs细胞和细胞培养:取左侧骨标本置于含有1%双抗体的预先制备的PBS中。将分离的长骨切断,过滤骨髓冲洗液,移入离心管中,离心后弃去上清液,将下层细胞重悬,培养细胞,培养3 d后更换培养液,然后第2天更换液体。待培养的细胞生长至培养皿的80%后,将细胞进行传代培养。
1.2.5大鼠检测指标:双能X线吸收仪测定骨密度。ELISA检测大鼠血清中骨钙素(BGP)和总碱性磷酸酶(TALP)含量。
1.2.5.1骨密度测定:取右侧骨标本使用多聚甲醛固定,并放置在可变系数设置为<1.0%的仪器中,双能X线吸收仪测量股骨骨密度。肱骨密度测量完毕再次使用生理盐水纱布,锡纸包裹,用于后续Western blot检测。
1.2.5.2ELISA:取保存于冰箱中的血液样本,按照ELISA试剂盒的说明,将样品和标准品分别装入平板中,加入生物素化抗体工作液和酶结合物工作液,并清洗平板。使用分光光度计检测450 nm处的A值,进而检测各组大鼠血清中BGP和TALP含量。
1.2.6细胞检测指标:qRT-PCR检测BMSCs细胞中miR-494和TLR4 mRNA的表达。Western blot检测大鼠BMSCs细胞中TLR4蛋白的表达。双荧光素酶报告基因验证miR-494与TLR-4的靶向关系。CCK8实验检测各组BMSCs的增殖能力。茜素红S染色检测各组BMSCs成骨分化情况。油红O染色检测各组BMSCs成脂分化情况(图1)。
1.2.6.1qRT-PCR:提取各组大鼠BMSCs细胞RNA,使用反转录试剂盒将RNA反转成cDNA。使用20 μL反应体系进行qRT-PCR。96 ℃预变性30 s,96 ℃变性10 s,60 ℃退火30 s,40个循环。TLR4 mRNA的表达以β-actin为内参,miR-494的表达以U6为内参计算相关基因的相对表达水平。
1.2.6.2Western blot:取各组大鼠右侧股骨标本,加入蛋白裂解液,在冰上浸泡1 h,然后在3 000 r/min下离心10 min。使用BCA测定蛋白质浓度。通过SDS-PAGE对蛋白质进行变性和分离。将蛋白质转移到聚偏二氟乙烯(PVDF)膜上,脱脂牛奶封闭2 h,分别使用一抗和二抗进行孵育。冲洗后,通过与化学发光试剂反应1 min,在黑暗中充分显影。以β-actin为内参。
1.2.6.3双荧光素酶报告基因验证miR-494与TLR-4的靶向关系:构建野生型WT-TLR-4和突变型MUT-TLR-4的3’UTR荧光素酶报告载体。利用LipofectamineTM 2000将miR-494和miR-control分别与WT-TLR-4和MUT-TLR-4共转染BMSCs细胞,分为以下4组:miR-control和WT-TLR-4共转染组、miR-494和WT-TLR-4共转染组、miR-control和MUT-TLR-4共转染组、miR-494和MUT-TLR-4共转染组。将各组BMSCs置于细胞培养箱内培养48 h,使用双荧光素酶报告检测试剂盒,检测各组细胞相对荧光素酶活性。
1.2.6.4CCK8实验检测各组BMSCs的增殖能力:将各组BMSCs按5×103个/孔,接种到96孔板,在37 ℃,5%CO2条件下孵育过夜。待细胞铺板孵育过夜后,弃去原培养基,对不同组细胞进行转染后,继续培养48 h,弃所有培养基,加入100 μL含有10% CCK8溶液的培养基,于37 ℃,5%CO2条件下继续培养3 h。使用分光光度计测定A450吸光度值,计算细胞增殖活性。
1.2.6.5茜素红S染色检测各组BMSCs成骨分化情况:收集各组BMSCs,以1×l05个/孔 接种于6孔板,使用茜素红S染色,根据倒置显微镜下观察结果,计数矿化结节数。评价标准:茜素红染色后钙结节呈橘红色;一般情况下,正常组钙结节数量多,骨质疏松时钙结节数量少。
1.2.6.6油红O染色检测各组BMSCs成脂分化情况:收集各组BMSCs,以l×l05个/孔接种于6孔板,使用油红O染色,在倒置显微镜下观察结果,计数脂肪细胞数。
1.3 统计学分析

2 结果
2.1 各组大鼠骨密度、BGP和TALP含量检测情况
检测各组大鼠骨密度和血清中BGP和TALP含量。和NC组大鼠相比,OP组大鼠骨密度和BGP含量显著降低,TALP含量显著升高,差异具有统计学意义(P<0.05)。和OP组大鼠相比,OP+miR-494 inhibitor组大鼠骨密度和BGP含量显著升高,TALP含量显著降低,差异具有统计学意义(P<0.05)。和OP+miR-494 inhibitor组大鼠相比,OP+miR-494 inhibitor+TLR-4 vector组大鼠骨密度和BGP含量明显降低,TALP含量显著升高,OP+miR-494 inhibitor+sh TLR-4组大鼠骨密度和BGP含量明显升高,TALP含量显著降低,差异具有统计学意义(P<0.05)(表1)。
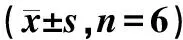
表1 各组大鼠血清中BGP和TALP含量比较Table 1 Comparison of BGP and TALP contents in serum of rats in each
2.2 各组大鼠BMSCs中miR-494和TLR4的表达情况
qRT-PCR检测各组大鼠BMSCs中miR-494和TLR4 mRNA的表达情况,Western blot检测各组大鼠股骨组织中TLR4蛋白的表达情况。和NC组大鼠相比,OP组miR-494、TLR4 mRNA和TLR4蛋白的表达显著升高(P<0.05)。和OP组大鼠相比,OP+miR-494 inhibitor组miR-494、TLR4 mRNA和TLR4蛋白的表达显著降低(P<0.05)。和OP+miR-494 inhibitor组大鼠相比,OP+miR-494 inhibitor+TLR-4 vector组TLR4 mRNA和TLR4蛋白的表达显著升高;OP+miR-494 inhibitor+sh TLR-4组TLR4 mRNA和TLR4蛋白的表达显著降低(P<0.05)。具体结果见表2。
表2 各组大鼠BMSCs中miR-494和TLR4 mRNA的表达水平比较Table 2 Comparison of the expression levels of miR-494 and TLR4 mRNA in BMSCs of rats in different
2.3 miR-494和TLR4的靶向关系验证
根据双荧光素酶活性检测结果显示,miR-494和WT-TLR-4共转染组(3.64±0.53)与miR-control和WT-TLR-4共转染组(1.09±0.21)、miR-control和MUT-TLR-4共转染组(1.13±0.26)、miR-494和MUT-TLR-4共转染组(1.06±0.22)相比,荧光素酶活性显著升高,差异具有统计学意义(P<0.05),而其他3组之间比较,荧光素酶活性无明显变化(P>0.05)。
2.4 各组大鼠BMSCs的增殖能力比较
CCK8检测各组大鼠BMSCs的增殖能力。结果显示,和NC组相比,OP组细胞增殖能力显著降低(P<0.05)。和OP组相比,OP+miR-494 inhibitor组细胞增殖能力显著升高(P<0.05)。和OP+miR-494 inhibitor组相比,OP+miR-494 inhibitor+TLR-4 vector组细胞增殖能力显著降低,OP+miR-494 inhibitor+sh TLR-4组细胞增殖能力显著升高(P<0.05),具体结果见表3。
表3 各组大鼠BMSCs的增殖情况对比Table 3 Comparison of BMSCs proliferation in different groups of
2.5 各组大鼠BMSCs成骨分化和成脂分化比较
和NC组相比,OP组细胞矿化结节数显著减少,脂肪细胞数显著升高(P<0.05)。和OP组相比,OP+miR-494 inhibitor组细胞矿化结节数显著升高,脂肪细胞数显著减少(P<0.05)。和OP+miR-494 inhibitor组相比,OP+miR-494 inhibitor+TLR-4 vector组细胞矿化结节数显著减少,脂肪细胞数显著升高,OP+miR-494 inhibitor+sh TLR-4组细胞矿化结节数显著升高,脂肪细胞数显著减少(P<0.05),具体结果见表4。
表4 细胞矿化结节数和脂肪细胞数对比Table 4 Comparison of the number of mineralized nodules and
3 讨论
骨质疏松症现在被认为是老年人的常见疾病,尤其是绝经后的老年妇女。卵巢功能退化引起的内分泌紊乱影响骨代谢,导致骨脆性增加,并随着时间的推移破坏骨组织的内部结构。骨组织脆性的增加会导致受损骨骨折,进而导致畸形和功能丧失,严重时会影响心脏和肺功能。尤其是由于骨质疏松引起的骨折和畸形,住院时间延长,严重影响患者的生活质量。
microRNA可以改变破骨细胞和成骨细胞的活性。几项临床和临床前研究发现,microRNA通过影响骨吸收、破骨细胞生成活性和骨形成来改变骨重建过程。miR-494作为microRNA家族成员之一,目前在恶性肿瘤中被广泛研究。在本研究中,qRT-PCR检测显示在骨质疏松大鼠中miR-494的表达显著升高。miR-494 inhibitor处理后,大鼠中miR-494表达显著降低,说明miR-494 inhibitor处理大鼠,成功使骨质疏松大鼠中miR-494表达受到抑制。miR-494被抑制后,大鼠骨密度显著升高,血清中骨密度指标BGP含量显著升高,TALP含量显著降低,大鼠骨质疏松程度明显改善。BGP是维持骨组织正常钙化的必需因子。TALP含量的升高可反应骨质疏松程度。以上结果证实miR-494在骨质疏松中存在表达差异,并可能通过影响大鼠骨形成,降低骨密度,进而加重骨质疏松。通过分离培养各组大鼠BMSCs,CCK8检测结果显示miR-494 inhibitor处理的大鼠BMSCs增殖能力显著升高。同时细胞矿化结节数显著升高,脂肪细胞数显著减少,这表明miR-494可能是通过抑制骨髓间充质干细胞增殖和成骨分化,促进骨髓间充质干细胞成脂分化,进而影响大鼠骨形成,降低骨密度,进而加重骨质疏松。
目前的研究表明,Toll样受体-4(TLR4)通路和骨质疏松之间存在密切的联系[9]。TLR-4信号通路可能通过影响下游转录因子的表达,进而影响骨形成相关蛋白的表达,并进一步抑制成骨细胞的作用。同时TLR-4信号通路可以通过抑制成骨相关标志物的表达,导致成骨细胞增殖受到抑制,导致骨细胞中矿化结节数形成减少,从而抑制成骨细胞的矿化[10-11]。先前已通过初步探究miR-494可能和TLR-4之间存在调控关系[8]。本研究通过双荧光素酶实验报告显示仅miR-494和WT-TLR-4共转染的细胞相对荧光素酶活性显著升高,这表明miR-494对相对荧光素酶活性的影响是通过促进TLR-4表达实现的,因此说明miR-494和TLR-4之间存在靶向关系。对骨质疏松大鼠进行miR-494 inhibitor和TLR-4 vector共处理,和miR-494 inhibitor单独处理相比,大鼠骨质疏松程度明显加重,细胞矿化结节数显著减少,脂肪细胞数显著增多,成骨分化被明显抑制,成脂分化显著增多。miR-494 inhibitor和shTLR-4共处理和miR-494 inhibitor单独处理的大鼠相比,可发现大鼠骨质疏松程度进一步改善,细胞矿化结节数显著增多,脂肪细胞数显著减少,成骨分化抑制明显减轻,成脂分化显著减少。以上结果表明,miR-494可能通过促进TLR4通路的表达,抑制骨髓间充质干细胞增殖和成骨分化,促进骨髓间充质干细胞成脂分化,进而影响大鼠骨形成,降低骨密度,进而加重骨质疏松。
本研究通过构建骨质疏松大鼠模型并提取大鼠骨髓间充质干细胞,经过体内实验探究miR-494在骨质疏松中的表达情况和调控机制,进一步完善骨质疏松的发病机制,并为临床辅助诊断和治疗骨质疏松提供新的理论依据。本研究主要的不足之处包括未更深入探究miR-494在骨质疏松中的调控网络,进一步完善miR-494的调控机制。
