甲基化转移酶样蛋白3介导的m6A修饰参与高静水压诱导的心房肌细胞电重塑
2023-12-12刘攀月肖海茵肖菲菲邝素娟邓春玉
刘攀月,曾 珑,肖海茵,肖菲菲,朱 蕊,杨 慧,邝素娟,邓春玉,饶 芳,魏 薇
(1.华南理工大学医学院,广东 广州 510006;2.广东省心血管病研究所心内科,广东省人民医院,广东省医学科学院,广东 广州 510080)
作为最常见的快速性心律失常之一,心房颤动(房颤,atrial fibrillation,AF)患者心脏衰竭、中风和外周血栓栓塞的风险增加。高血压(hypertension,HTN)是AF发生的独立危险因素,流行病学研究显示,HTN使新发AF的风险增加1.8倍,使永久性AF的发病风险增加1.5倍[1]。HTN患者由于高静水压、牵张力和神经激素等影响,左心房发生弥漫性电重塑,使AF易于发生。电重塑是指由心房肌细胞中Ca2+或者K+通道密度改变导致动作电位时程(action potential duration,APD)缩短,折返的发生和电特性的改变,其中L型钙通道蛋白(L-type calcium channel α1C,Cav1.2)的减少及L型钙电流(L-type calcium current,ICa,L)的减少是APD缩短的主要原因[2]。前期研究表明,高静水压下HL-1细胞发生ICa,L和超速延迟整流钾电流(Ikur)改变,AF易感性增加[3-4],但高血压致AF的具体分子机制尚不清楚。
转录后修饰可调控细胞新陈代谢、细胞增殖和细胞迁移等,与心血管疾病密切相关[5]。m6A修饰属于转录后修饰的一种类型,由各种转甲基化酶(METTL3、METTL14、RBM15、RBM15B、KIAA1429、WTAP等)和去甲基化酶(FTO和ALKBH5)动态调控。转甲基化酶或去甲基化酶与靶RNA结合后,可通过添加/去除m6A修饰调节RNA的生理过程,如稳定性、定位、剪接、翻译等生理过程[6]。研究表明,m6A修饰可间接参与心肌细胞电活动,冠状动脉结扎诱导心梗小鼠模型METTL3明显升高,m6A修饰小胶质细胞中Toll样受体4 mRNA,从而增强交感神经活动诱导室性心律失常的发生[7];m6A修饰可直接参与心肌细胞电活动,敲除FTO的小鼠发生心脏电学重塑和结构重塑,心率变异性增加、心室复极化时程改变,表明m6A甲基化的增加可以使小鼠更容易发生快速性心律失常[8]。然而,依赖于METTL3的m6A修饰是否在HTN致AF的电生理机制中发挥表观遗传作用还有待确定。我们拟通过本研究探讨METTL3介导的m6A修饰是否参与高静水压对ICa,L的调控,进而促进房颤的发生发展。
1 材料与方法
1.1 主要试剂与仪器抗m6A抗体(Synaptic Systems,202003);抗METTL3抗体(Abcam,ab195352);抗Cav1.2抗体(Alomone,ACC003AN7108);抗GAPDH抗体(Proteintech,60004-1-I);兔二抗(Jackson ImmunoResearch,111-035-003);鼠二抗(Jackson ImmunoResearch,111-015-003);STM2457(MedChemExpress,2499663-01-1);亚甲蓝(BioReagent,M4159-25G);反转录试剂盒(Accurate Biotechnology,AG11734);预混型qPCR试剂盒(Accurate Biotechnology,AG11733);荧光定量PCR仪(qTOWER3 G);RIP试剂盒(BersinBio,Bes5101);紫外交联仪(ZIXING1793);正置荧光显微镜(Nikon,80i);EPC10膜片钳放大器及附件(HEKA)。
1.2 方法
1.2.1实验动物 SPF级C57BL/6J雄性小鼠(35 g左右),使用Alzet微渗透泵持续皮下泵入血管紧张素4周(给药剂量为1 000 ng·kg-1·min-1),构建HTN模型小鼠(n=6)。选取另外6只性别、体质量匹配的C57BL/6J小鼠作为对照,动物从湖南斯莱克景达实验动物有限公司购买,许可证号为SCXK(湘)2019-0004。小鼠模型造模成功后留取心房组织,分装后放入液氮速冻再保存至-80 ℃冰箱。本研究已经通过广东省人民医院(广东省医学科学院)动物实验伦理委员会审查(No 201904010451)。
1.2.2细胞培养与干预 本研究所使用的小鼠心房肌细胞系HL-1由美国路易斯安那州立大学William Claycomb教授慷慨赠送。Claycomb专用培养基、10%胎牛血清、0.1 mmol·L-1去甲肾上腺素和2 mmol·L-1左旋-谷氨酰胺培养细胞。待细胞密度长至约80%时,使用0.05%胰酶消化后将细胞种于3.5 mm×10 mm小皿中进行后续干预实验。以常压(0 mmHg)作为对照,设置了不同压力梯度(20、40 mmHg)干预细胞48 h。为了观察干预METTL3能否逆转高静水压下Cav1.2的下调,分别利用 STM2457 和si-METTL3来干预METTL3的功能和合成,设置对照组、高压+DMSO组、高压+STM2457组、高压+阴性对照si-ctrl组及高压+si-METTL3组。STM2457 配制浓度1、3、10 μmol·L-1,阴性对照si-ctrl和si-METTL3(CAAGGAAGAGTGCATGAAA)配制成50 nmol·L-1分别转染HL-1细胞。
1.2.3斑点印迹实验 提取组织或细胞中RNA后检测浓度,将一定量的RNA滴于N+尼龙膜上,自然风干尼龙膜后置于紫外交联仪下照射10 min,5%脱脂牛奶进行室温封闭2 h,1 ∶1 000稀释的抗m6A抗体过夜孵育N+尼龙膜。随后兔二抗(1 ∶5 000)室温孵育1 h,PBST 5 min洗3次后置于化学发光成像仪下显影。显影后0.01%亚甲蓝孵育N+尼龙膜10 min,在自然光下拍摄尼龙膜作为对照。
1.2.4mRNA定量检测和蛋白定量检测 qRT-PCR和Western blot详细实验步骤见既往发表的文献[9],各引物序列见Tab 1。

Tab 1 Sequences of primers used for real RT-PCR
1.2.5全细胞膜片钳实验 细胞干预充足时间后使用预热的0.05%胰酶消化3 min,显微镜下可见细胞轮廓变圆终止消化,30 min后进行全细胞膜片钳实验。使用进液针将电极内液注入玻璃电极后,要求入液时电极电阻达到2~5 MΩ。选取大小合适的细胞,电极尖端轻触细胞表面可产生微小形变,给予些许负压后可观察电阻变大,待电阻达到1 GΩ(高阻封接)时使用电刺激破膜,补偿快慢电容、串联电阻。全细胞状态稳定后,电压钳模式记录ICa,L及膜电容(Cm),电流钳模式记录动作电位(AP),最后使用Clampfit 10.4测量数据及统计。HL-1 ICa,L电极内液成分:CsCL 100 mmol·L-1,TEA-CL 20 mmol·L-1,EGTA 10 mmol·L-1,HEPES 10 mmol·L-1,Tris将pH调至7.2;ICa,L细胞外液成分:TEA-CL 140 mmol·L-1,CaCl2·2H2O 5 mmol·L-1,MgCl2·6H2O 2 mmol·L-1,HEPES 10 mmol·L-1,Glucose·H2O 10 mmol·L-1,Tris将pH调至7.4;HL-1 AP电极内液成分:KCl 140 mmol·L-1,MgCl2·6H2O 1 mmol·L-1,HEPES 10 mmol·L-1,EGTA 5 mmol·L-1,KOH将pH调至7.2;AP细胞外液成分:NaCl 136 mmol·L-1,KCl 5.4 mmol·L-1,MgCl2·6H2O 1.0 mmol·L-1,NaH2PO4·2H2O 0.33 mmol·L-1,HEPES 238.3 mmol·L-1,D-Glucose 10 mmol·L-1,CaCl2·2H2O 1.8 mmol·L-1,NaOH将pH调至7.4。
1.2.6RNA结合蛋白免疫沉淀(RIP)实验 使用BersinBio RIP试剂盒(BersinBio,Bes5101)进行RIP实验,根据试剂的说明书,使用裂解缓冲液收集细胞,并将裂解后的RNA-蛋白质复合物与转载IgG或抗METTL3抗体(Abcam,ab195352,1 ∶100)的磁珠4 ℃过夜孵育,RIP洗涤缓冲液将磁珠-蛋白-RNA复合物于4 ℃摇床上洗涤10 min,然后用RIP裂解缓冲液于4 ℃摇床上洗涤5 min,收集洗脱下来的含有RNA的上清,并用TRIzol/氯仿/异丙醇提取RNA,利用qRT-PCR检测RNA的表达量。

2 结果
2.1 HTN小鼠左心房组织RNA m6A表达增加及形态学改变斑点印迹实验结果显示与对照组野生型(WT)小鼠相比,HTN组小鼠左心房组织中m6A含量增加(定量为400 ng时,P<0.01;定量为200 ng时,P<0.05)(Fig 1A-B);相较于WT组小鼠,HE染色提示,HTN组小鼠左心房心肌细胞结构更为紊乱,同时间质含量增加,Masson染色提示HTN小鼠左心房组织胶原含量增加(Fig 1C)。m6A的含量是否与HTN小鼠发生心房重塑有关还需进一步探究。
2.2 HTN小鼠左心房组织METTL3,Cav1.2的表达减少RNA m6A水平主要是由转甲基化酶和去甲基化酶介导,因此我们进一步检测了WT和HTN组小鼠左心房组织中,转甲基化酶METTL3、METTL14、RBM15、RBM15B、KIAA1429、WTAP和去甲基化酶FTO、ALKBH5的RNA表达量。结果发现,与WT组相比,HTN组小鼠心房组织中METTL3明显增加(P<0.01),而FTO、KIAA1429则明显下调(P<0.05)(Fig 1D)。我们进一步在蛋白水平验证METTL3的表达改变,发现结果与mRNA水平一致(P<0.01),同时Cav1.2在HTN组中明显减少(P<0.01)(Fig 1E-F)。Cav1.2的减少是否与METTL3介导的m6A修饰有关需进一步验证。
2.3 高静水压对HL-1细胞RNA m6A、METTL3、Cav1.2的影响体外予以不同的静水压(20 mmHg和40 mmHg)模拟心房高压的病理状态,斑点印迹实验显示HL-1中m6A的含量随着静水压的增加而增加(P<0.01 in 20 mmHg,P<0.01 in 40 mmHg)(Fig 2A,B),同时HL-1细胞中METTL3蛋白的含量随着静水压的升高而增加(P<0.01 in 20 mmHg,P<0.01 in 40 mmHg),而Cav1.2蛋白表达则随着静水压的升高而降低(P<0.05 in 20 mmHg,P<0.01 in 40 mmHg)(Fig 2C,D)。提示高静水压下心房肌细胞RNA m6A、METTL3增加,同时细胞中Cav1.2减少,与动物模型实验结果一致。
2.4 METTL3抑制剂STM2457逆转高静水压所致的HL-1细胞Cav1.2下调为了进一步明确METTL3在高静水压所致心房肌细胞的Cav1.2表达和ICa,L下降中的作用,HL-1细胞加压之前,予METTL3的特异性抑制剂 STM2457(1、3、10 μmol·L-1)预处理细胞3 h。斑点印迹实验结果表明,STM2457 对高静水压组的HL-1细胞具有m6A抑制作用(P<0.01 in 3,10 μmol·L-1),而相较于加压组,STM2457使Cav1.2的表达量有所增加,并在10 μmol·L-1时差异有统计学意义(P<0.05 in 10 μmol·L-1)(Fig 3A-C)。
同时使用全细胞膜片钳技术记录常压下、高静水压下、高静水压+STM2457(10 μmol·L-1)各组HL-1细胞ICa,L和动作电位(AP)。对比常压下,使用40 mmHg高静水压处理HL-1细胞时,细胞APD50、APD70、APD90均明显缩短,而使用STM2457处理高静水压下的HL-1细胞时,则APD50、APD70、APD90均有增加(P<0.05)(Fig 3D,F),但动作电位振幅(action potential amplitude,APA)差异无显著性(Fig 3E)。ICa,L峰值电流密度在高静水压组明显降低,并在10 μmol·L-1STM2457 的干预下逆转(峰值电压为+10 mV时,2.70±0.26 pA/pF in 0 mmHg+DMSO groupvs-1.61±0.28 pA/pF in 40 mmHg+DMSO groupvs2.25±0.45 pA/pF in 40 mmHg±STM2457 ,P<0.01)(Fig 3G-H)。各组ICa,L的激活、失活电压-电流关系,复活时间曲线无明显差异(P>0.05)(Fig 3I-J)。
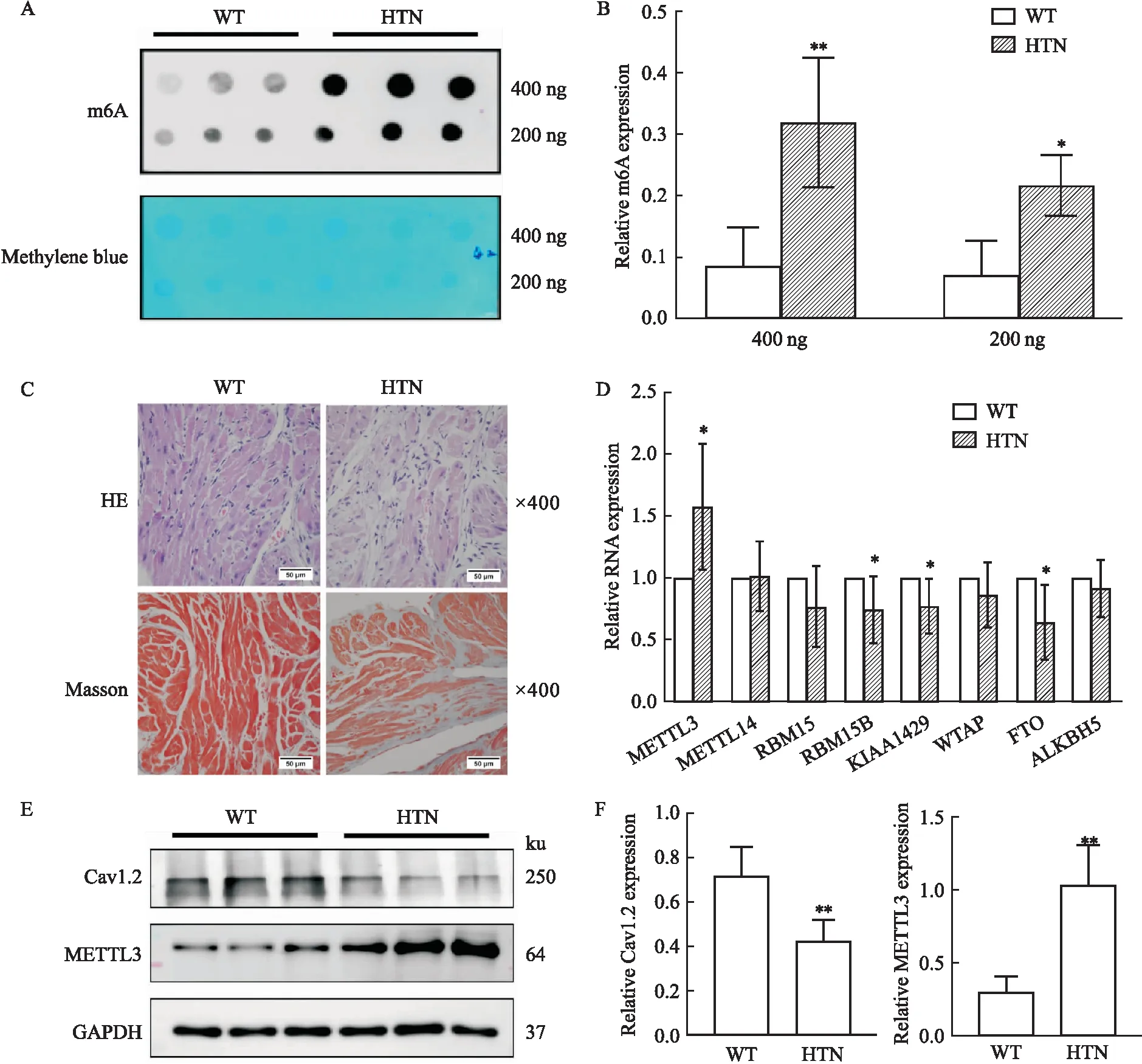
Fig 1 Expression of m6A-modified mRNA,methylases,Cav1.2,METTL3 in left atrial

Fig 2 Effect of high hydrostatic pressure on m6A-modified mRNA,METTL3 and Cav1.2 in HL-1
2.5 敲低METTL3可逆转高静水压所致的HL-1细胞Cav1.2下调敲低METTL3后观察对高静水压所致心房肌细胞电重塑的逆转作用。予METTL3 siRNAs转染HL-1细胞后,加压干预48 h(40 mmHg)。结果发现,成功敲低METTL3后,HL-1细胞的RNA m6A表达水平减少(P<0.01)(Fig 4A-B),说明METTL3发挥了转甲基酶的作用。Western blot结果显示,敲低METTL3,可使Cav1.2较对照组有增加(P<0.01)(Fig 4D-F)。为了验证METTL3对Cav1.2的调控作用,我们使用RIP及qRT-PCR检测HL-1中 METTL3是否与CACNA1C结合。结果显示,相对于IgG阴性对照,使用METTL3免疫沉淀后结合的CACNA1C明显增加(P<0.01)(Fig 4C),说明METTL3可以直接结合CACNA1C,从而调控其蛋白Cav1.2的表达,影响心房肌细胞电重塑。
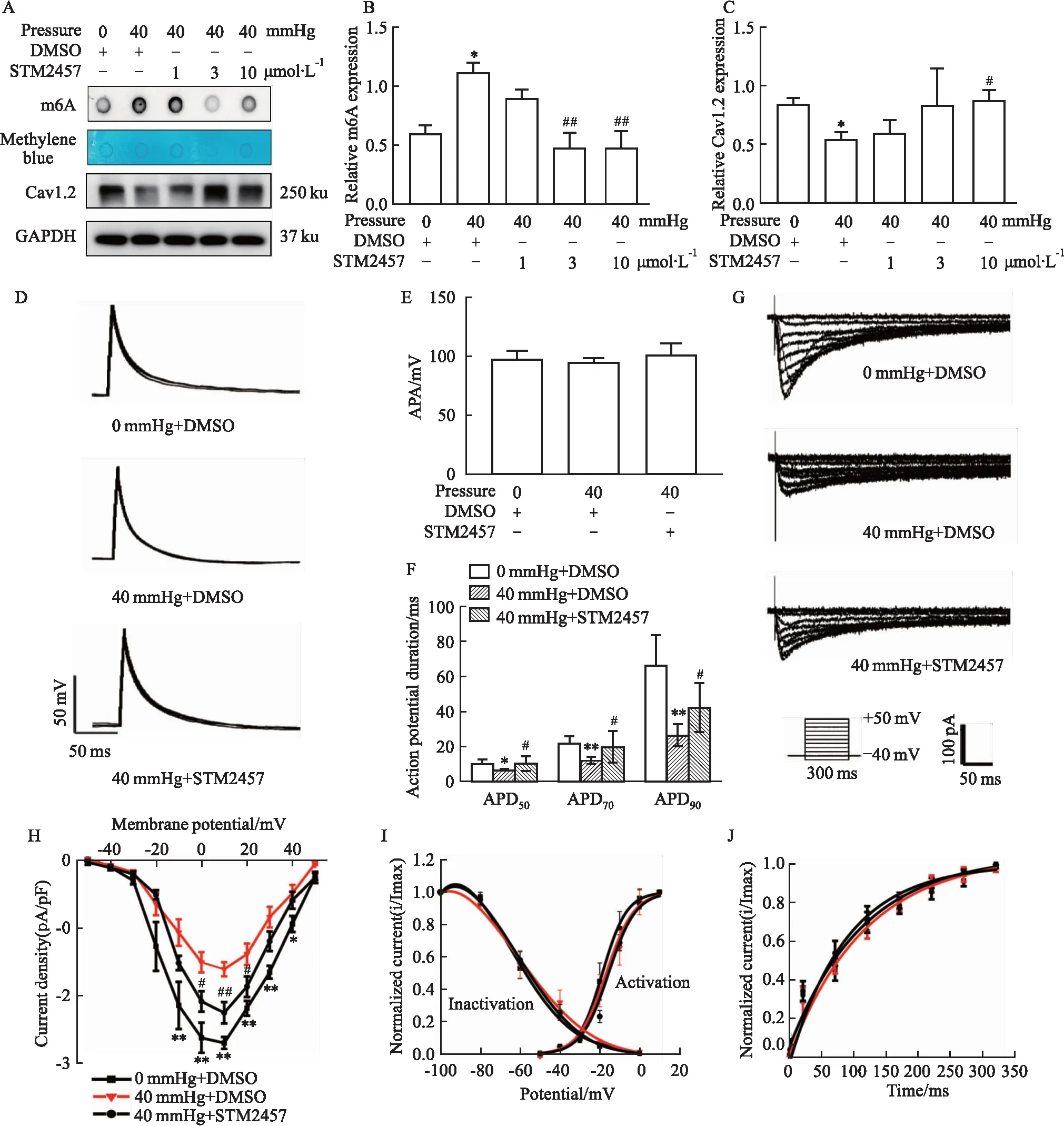
Fig 3 Effect of STM2457 on m6A-modified mRNA,METTL3 and iron channel current

Fig 4 Effect of si-METTL3 in HL-1 cells after treated with hydrostatic
3 讨论
研究表明:(1)与对照组相比,HTN小鼠心房间质纤维化增加,心肌细胞分布紊乱,离子通道蛋白Cav1.2表达减少,提示高压下诱导心房肌细胞发生电重塑。同时小鼠心房内m6A表达量增加,METTL3蛋白表达升高。(2)随着静水压的升高,心房肌细胞HL-1细胞Cav1.2蛋白表达减少,m6A和METTL3蛋白表达增加;抑制METTL3的功能与合成可明显降低高静水压下HL-1细胞 m6A的表达以及改善HL-1细胞的电重构,同时证明了METTL3直接结合CACNA1C,从而发生转甲基修饰引起心房肌细胞电重塑。
心房肌细胞电重塑是AF发生和维持的重要机制,特别是L型钙通道蛋白表达的降低和ICa,L的减少造成APD缩短是AF电重塑的主要内容,既往研究表明,在AF患者中Cav1.2的表达明显减少,缩短了AF及患者的APD,从而提高AF的诱发率和促进AF持续发生[10];快速起搏心房兔模型Cav1.2泛素化降解增加,缩短APD和减少ICa,L,促进心房电重构[11]。HTN可通过多种机制诱导心房肌细胞电重塑,包括改变离子通道表达、影响Ca2+的内流和外流以及影响细胞内Ca2+稳态等[12]。L型钙通道在HTN诱导的心房肌细胞电重塑中发挥着重要作用,高静水压直接激活Piezo1,将机械力转变为生物信号诱导心房肌细胞ICa,L的电生理重塑[13];高静水压培养下,心房肌细胞的肾素-血管紧张素系统被激活,由FAK-Src促进ICa,L减小,AF易感性增加[4],与之前的研究结果一致,我们的研究结果表明,在HTN小鼠模型中Cav1.2蛋白表达明显减少,提示HTN小鼠心房肌细胞L型钙通道发生改变。同时HL-1施予高静水压(40 mmHg),APD50、APD70、APD90均明显缩短,提示高静水压下心房肌细胞发生电重塑。有趣的是,高静水压下HL-1细胞L型钙通道的激活、失活动力学并未发生改变,说明ICa,L密度的降低主要由Cav1.2蛋白表达减少引起。
上述研究[4,14]都集中在转录水平调控高静水压下心房肌细胞电重塑,尚不清楚在此过程中L型钙通道的表达和功能变化是否与转录后修饰有关。近年来,RNA的m6A甲基化修饰作为一种转录后修饰日益受到关注,在m6A转甲基酶复合物中,METTL3是催化m6A甲基化的主要亚基,负责在RNA上安装甲基,细胞失去动态平衡时,METTL3在细胞内触发全局mRNA甲基化,通过调节不同的mRNA亚群来决定其功能和命运[14]。多项研究表明,血管紧张素II通过METTL3诱导心肌细胞纤维化、心肌肥厚[15];血管紧张素II以浓度依赖的方式降低HTN豚鼠心室肌细胞ICa,L峰值密度[16]。但是METTL3是否参与高压下Cav1.2蛋白表达下降,目前尚无相关报道。因此,本研究将m6A与AF联系起来,在高压的作用下,无论是小鼠体内还是心房肌细胞HL-1细胞中,m6A和METTL3均有增加,Cav1.2蛋白表达量减少;在体外使用 STM2457 处理高静水压下的HL-1细胞后,逆转了高静水压下ICa,L的降低和APD50、APD70、APD90的缩短。同时RIP实验证明了METTL3与CACNA1C直接结合,表明METTL3直接参与了高静水压下CACNA1C的m6A修饰,进而引起L型钙通道蛋白表达减少,参与心房肌细胞电重塑,使AF易于发生。潘振伟教授实验团队使用METTL3基因敲除小鼠来探索m6A修饰在心脏电重塑中的效应,敲除METTL3可通过减少小鼠中CACNA1C的 m6A 修饰来上调 Cav1.2的表达,延长APD,从而增加室性心律失常的易感性[17]。我们研究结果与潘振伟教授团队不一致可能是因为采用了不同的小鼠模型(我们的是血管紧张素诱导的HTN模型小鼠,他们的是METTL3基因敲除杂合子小鼠)及不同的研究部位(我们研究心房,他们研究心室),但都证明了METTL3对CACNA1C具有m6A修饰作用,METTL3可直接参与心肌细胞电活动的调控。
STM2457 是METTL3-METTL14转甲基酶复合物的有效抑制剂,其抑制作用为竞争性结合METTL3的甲基结合位点,却并未破坏METTL3-METTL14复合物的蛋白结构。且 STM2457 对METTL3具有高度的选择性,对其他RNA甲基转移酶没有抑制作用[18]。基于上述研究,我们使用了 STM2457 抑制高静水压下HL-1中METTL3的转甲基作用,逆转了ICa,L的下调,但并未在HTN模型小鼠中使用 STM2457,因此 STM2457 对HTN小鼠心房肌细胞的影响是否也像体外实验一样降低AF易感性仍需进一步探究。
然而,本研究具有以下局限性有待完善:(1)只研究了一种离子通道电流(ICa,L),而致AF易感性增加的还有其他离子通道电流,如Ikur,Ik1等;(2)未在体内探究干预METTL3对HTN小鼠的影响,同时STM2457 在体内是否会对其他细胞产生毒性效应需进一步探索。
综上所述,本研究证实了METTL3是高静水压致AF的关键调节因子,METTL3通过m6A修饰CACNA1C参与高静水压诱导的ICa,L下降,引起心房肌细胞电重塑;抑制METTL3可逆转高静水压所致的Cav1.2下调,缓解高静水压诱导心房肌细胞膜离子通道蛋白重塑,从而降低AF易感性。本研究为RNA表观遗传学参与高静水压致AF的发病机制提供了实验证据,为HTN合并AF的治疗提供了潜在的途径。
