野黄芩素通过促进胆固醇外流途径抑制巨噬细胞泡沫化的作用机制
2023-12-12吴若琳黄裕鸿王天琦孙文龙李敬达
吴若琳, 黄裕鸿, 王天琦, 孙文龙, 李敬达
(长江大学 1.生命科学学院、 2.农学院,湖北 荆州 434000;3. 山东理工大学生命与医药学院,山东 淄博 255000)
动脉粥样硬化(atherosclerosis,AS)是一种以胆固醇在动脉血管内膜过度沉积为主要病理特征的慢性血管类疾病,因其引发的血栓形成及动脉血管狭窄可直接导致冠心病、心肌梗死以及缺血性脑卒中等疾病的发生,故AS已成为威胁人类生命健康的重大疾病之一,探究其治疗方法和新型药物对于改善人类健康具有重要意义。大量研究表明,氧化低密度脂蛋白(oxidized low density lipoprotein,oxLDL)所诱导的巨噬细胞泡沫化是AS发病过程中的关键起始环节[1]。在AS早期阶段,巨噬细胞可通过其表面清道夫受体,大量摄取血管内膜组织中的oxLDL,并以胆固醇酯的形式存储于细胞中[2]。另一方面,巨噬细胞还可通过三磷酸腺苷结合盒转运体A1(ATP-binding cassette transporter A1,ABCA1)介导的胆固醇外流途径,将胆固醇酯经载脂蛋白A1(apolipoprotein A-1,ApoA-1)及高密度脂蛋白外排至细胞外,以维持细胞内胆固醇的动态平衡[3]。然而,随着oxLDL被巨噬细胞不断摄入,胆固醇流入速率大于胆固醇外流速率,致使胆固醇代谢平衡被打破,造成胆固醇在细胞中过量积累,进而引发巨噬细胞泡沫化,最终导致AS斑块形成并推动病情发展[2]。因此,抑制巨噬细胞泡沫化对改善AS意义重大。
目前,天然产物已成为新药物先导结构的重要来源,并展现出较高的药物开发价值。野黄芩素(Scutellarein)是一种黄酮类化合物,主要存在于黄芩、木蝴蝶、半枝莲等多种中药材中[4-5]。近年来研究发现,野黄芩素除了具有良好的生物利用度和抗氧化活性外,还在治疗老年性痴呆症、心脑血管疾病、中风后遗症和癌症等方面有明显的疗效[6]。既往研究发现,野黄芩素不仅具有较高的抗氧化活性[7],还可在LPS刺激的RAW264.7巨噬细胞中,通过阻断NF-κB和MAPKs通路发挥抗炎作用[8];此外,野黄芩素可通过抑制凝血酶及G蛋白偶联受体活性减少血小板的黏附和聚集[9],并能够阻止大鼠体内动脉或静脉血栓的形成[10]。以上这些研究都表明,野黄芩素具有潜在的抗AS作用,但其是否能够增强胆固醇外流途径并抑制巨噬细胞泡沫化目前尚不清楚。为此,利用50 mg·L-1oxLDL,建立了THP-1巨噬泡沫细胞模型,旨在分析并阐明野黄芩素抑制巨噬细胞泡沫化的作用效果及潜在机制,为今后野黄芩素用于AS的临床治疗提供重要的理论依据。
1 材料与方法
1.1 药物与试剂野黄芩素(批号:S0327)、NBD标记胆固醇(批号:810250P)、佛波醇12-十四酸酯13-乙酸酯(PMA)(批号:79345)和罗格列酮(批号:R2408)均购自美国默克Sigma公司;总胆固醇(total cholesterol,TC)含量检测试剂盒(批号:A110-1-1)购自南京建成生物工程研究所;抗His标签抗体(批号:105327-MM02T)、人源ApoA-1重组蛋白(批号:10686-H02H)及人源PPARγ重组蛋白(批号:12019-H20B)均购自北京义翘神州科技有限公司;RIPA裂解液(批号:P0013B)、Lipo8000转染试剂(批号:C0533)、双萤光素酶报告基因检测试剂盒(批号:PG027)、pRL-TK对照载体(批号:D2760)及ECL化学发光试剂盒(批号:P0018S)均购自上海碧云天生物技术有限公司;PPARγ-siRNA及Scramble对照siRNA购自上海吉玛制药技术有限公司;ABCA1抗体(批号:A7228)、PPARγ抗体(批号:A0270)、LXRα抗体(批号:A2141)、β-actin肌动蛋白抗体(批号:PKSH033261)及山羊抗兔二抗(批号:E-AB-1034)均购自武汉爱博泰克生物科技有限公司;PPRE报告基因载体(实验室前期构建)。
1.2 仪器倒置显微(德国徕卡公司);二氧化碳细胞培养箱(美国赛默飞世尔公司);多功能酶标仪(美国molecular devices公司);凝胶成像系统(美国伯乐公司);蛋白垂直电泳仪、蛋白转膜仪(北京六一生物科技公司);低速台式离心机(湖南湘仪实验室仪器开发有限公司);高速冷冻离心机(美国赛默飞世尔公司)。
1.3 方法
1.3.1THP-1细胞培养及巨噬化 人单核细胞株THP-1由武汉大学保藏中心提供。THP-1细胞解冻复苏,接种于RPMI 1640完全培养基中(含有10 mmol·L-14-羟乙基哌口秦乙磺酸、1 mmol·L-1丙酮酸钠、50 mmol·L-12-巯基乙醇、2 mmol·L-1谷氨酰胺、1×105IU·L-1青霉素、0.1 g·L-1链霉素和10%热灭活的胎牛血清),并置于37 ℃、5% CO2条件下悬浮培养。待细胞生长至70%密度时,1500 r·min-1离心5 min,收集细胞,并以1 ∶5的比例传代培养。 传代5次后,取对数生长期的细胞,使用含有PMA(160 nmol·L-1)的完全培养基处理24 h,以诱导细胞分化为巨噬细胞。
1.3.2MTT法检测THP-1细胞存活率 将THP-1细胞以每孔1×104个细胞的密度接种于96孔板中,置于37℃、5% CO2条件下培养过夜。PMA诱导细胞巨噬化后(参照“1.3.1”),分别使用0.1%的DMSO作为空白对照组,用浓度为1、2.5、5、10、20、40、80 μmol·L-1野黄芩素(野黄芩素组)刺激细胞24 h。PBS清洗细胞后,参照检测试剂盒使用说明,测定各组样品的细胞存活率。
1.3.3细胞内TC含量的测定 将THP-1细胞以每孔1×106个细胞的密度接种于6孔板中,PMA诱导细胞巨噬化后,细胞分为空白对照组(DMSO 0.1%)、oxLDL组(50 mg·L-1)、oxLDL+野黄芩素组(5、10和20 μmol·L-1),参照TC含量检测试剂盒使用说明,测定各组细胞的TC含量。
1.3.4THP-1巨噬细胞胆固醇外流速率的检测 将THP-1细胞以每孔2.5×105个细胞的密度接种于24孔板中,PMA诱导细胞巨噬化后,细胞分为空白对照组(DMSO 0.1%)和野黄芩素组(5、10、20 μmol· L-1)预处理24 h后,用PBS洗净细胞,使用含有0.2% BSA及50 mg· L-1ApoA-1的完全培养基孵育细胞6 h。分别收集细胞沉淀及上清液,并用RIPA裂解液裂解细胞沉淀。将上清液和细胞裂解液分别加入96孔黑板,并在469 nm激发波长及537 nm发射波长条件下,测定各组样品中NBD的荧光强度。最终,细胞胆固醇外流的速率=上清液荧光强度/(上清液荧光强度+细胞裂解液荧光强度)。
1.3.5分子对接分析PPARγ与野黄芩素及罗格列酮的结合作用 从Protein Data Bank数据库中下载PPARγ-LBD结构域的晶体结构(PDB ID: 5YCP),并利用ChemBio3D软件构建野黄芩素及罗格列酮的PDB结构。分别对PPARγ-LBD晶体结构、野黄芩素和罗格列酮的PDB结构进行电荷分配、加氢去水、能量最小化等预处理操作,并分别利用AutoDock 4.2.6软件包中的AutoGrid和Autodock模块生成30×30×30 Å大小的对接网格和按照1 ∶1 的比例进行半柔性对接模拟分析。在对接过程中,PPARγ-LBD的晶体结构被设定为刚性,野黄芩素和罗格列酮的PDB结构被设定为柔性,对接方法为Lamarchian遗传算法,其他参数均为默认值。对接结束后,利用PyMOL软件绘制结合自由能最低的对接构象模型。
1.3.6ELISA法检测PPARγ与野黄芩素及罗格列酮的结合活性 分为对照组(仅添加PBS)、罗格列酮组和野黄芩素组,将PBS、罗格列酮(5、10、20 μmol·L-1)及野黄芩素(5、10、20 μmol·L-1)以50 μL/孔分别加入96孔酶标板中,4 ℃条件下包被过夜后,使用1%明胶(100 μL/孔)封闭酶标板1 h,并于37 ℃条件下孵育PPARγ重组蛋白(20 mg·L-1, 50 μL/孔)1 h。随后,各孔先后孵育抗His标签抗体(1 ∶1 000)及HRP标记山羊抗兔二抗(1 ∶3 000),37 ℃条件下1 h,使用PBST(200 μL/孔)清洗酶标板4次。最后,用OPD显色,并在492 nm处读取吸光度值。
1.3.7双荧光素酶报告基因检测PPARγ的转录活性 将THP-1细胞以每孔2.5×105个细胞的密度接种于24孔板中,PMA诱导细胞巨噬化后,使用Lipo8000转染试剂将PPRE报告基因载体(800 ng)及pRL-TK对照载体(200 ng)共转染入细胞,具体方法参照Lipo8000转染试剂使用说明。转染36 h后,将细胞分为空白对照组(DMSO 0.1%)、罗格列酮组(5、10、20 μmol·L-1)和野黄芩素组(5、10、20 μmol· L-1),孵育24 h后,使用双荧光素酶报告基因检测试剂盒先后测定细胞中海肾及萤火虫荧光素酶的化学发光强度,并根据二者的比值计算各组细胞的PPARγ转录活性。
1.3.8Western blot检测细胞内蛋白表达水平 将THP-1细胞以每孔1×106个细胞的密度接种于6孔板,并放置于37 ℃、5% CO2条件下培养过夜。PMA诱导细胞巨噬化后,细胞如“1.3.3”中分组处理,孵育24 h,收集各组细胞并用RIPA裂解液裂解后,利用BCA蛋白浓度检测试剂盒测定裂解液中的蛋白浓度。取50 μg蛋白样品,加入上样缓冲液,并放置于沸水浴中变性5 min。随后,将各组蛋白样品进行12% SDS-PAGE电泳分离,并将蛋白转膜至PVDF膜。用5% 脱脂牛奶溶液封闭1 h后,将PVDF膜置于相关蛋白一抗稀释液(1 ∶1 000)中,4 ℃条件下孵育过夜。TBST溶液清洗膜3次后,使用HRP标记山羊抗兔二抗(1 ∶3 000),室温条件下孵育1 h。洗膜3次后,使用ECL化学发光试剂盒对膜进行显影和拍照分析。
1.3.9siRNA介导的PPARγ基因沉默 将THP-1细胞以每孔1×106个细胞的密度接种于6孔板。PMA诱导细胞巨噬化后,将细胞分为scramble对照组和PPARγ沉默组,使用Lipo8000转染试剂将scramble-siRNA(100 nmol·L-1)及PPARγ-siRNA(100 nmol·L-1)分别转染入细胞,转染24 h后,通过Western blot方法分析细胞模型中PPARγ的基因沉默效果。siRNA转染后,使用DMSO(1%,对照组)及野黄芩素(20 μmol·L-1,野黄芩素组)分别刺激scramble-siRNA组及PPARγ-siRNA组细胞24 h,并分析各组样品中LXRα、ABCA1蛋白表达及胆固醇外流速率的变化情况。此外,在研究PPARγ基因沉默对野黄芩素抑制巨噬细胞泡沫化的影响实验中,siRNA转染后,使用DMSO(1%)、oxLDL(50 mg· L-1)以及oxLDL(50 mg· L-1)与野黄芩素(20 μmol·L-1)的混合液分别刺激scramble-siRNA组及PPARγ-siRNA组细胞24 h,最后利用TC含量检测试剂盒测定各组细胞的TC含量。

2 结果
2.1 野黄芩素通过增强胆固醇外流途径抑制巨噬细胞泡沫化首先,通过MTT法分析不同浓度野黄芩素对THP-1巨噬细胞活力的影响。结果如Fig 1A所示,与空白对照组相比,1~20 μmol·L-1野黄芩素对THP-1巨噬细胞活力未产生明显的抑制作用。然而,当野黄芩素剂量达到40、80 μmol·L-1时,THP-1巨噬细胞活力被明显抑制。因此,5、10、20 μmol·L-1浓度的野黄芩素被认为是安全剂量,并被用于后续的药物刺激实验中。之后,本文分析了不同浓度野黄芩素对巨噬细胞胆固醇蓄积和外流作用的影响。结果如Fig 1B所示,与空白对照组相比,50 mg·L-1oxLDL处理24 h,可使细胞内胆固醇含量提高3.1倍,而野黄芩素的刺激作用可明显抑制oxLDL诱导的胆固醇蓄积,并且该抑制作用呈现明显的剂量依赖性。此外,野黄芩素还可通过剂量依赖的方式增强巨噬细胞的胆固醇外流速率(Fig 1C)。以上结果表明,野黄芩素可能通过加速胆固醇外流途径阻止巨噬细胞的泡沫化。
2.2 PPARγ与野黄芩素及罗格列酮结合作用分析随后,通过分子对接方法分析了野黄芩素及PPARγ化学合成型激活剂罗格列酮(分子结构如Fig 2A)与PPARγ-LBD结构域的结合情况。结果显示,野黄芩素及罗格列酮均可插入至PPARγ-LBD结构域的T形配体结合区域(橙色区域);罗格列酮能够与PPARγ的His-323及His-449形成两根氢键(Fig 2D),野黄芩素可与PPARγ的Ser-289、His-323、His-449及Tyr-473形成四根氢键(Fig 2F),说明野黄芩素能与PPARγ结合;同时罗格列酮及野黄芩素与PPARγ-LBD结构域的对接结合自由能分别为-6.32 kcal·mol-1(Fig 2E)和-7.21 kcal·mol-1(Fig 2G),表明与罗格列酮相比,野黄芩素具有更低的结合自由能。以上结果提示,野黄芩素可结合PPARγ,且与罗格列酮相比其相互作用更为稳定。
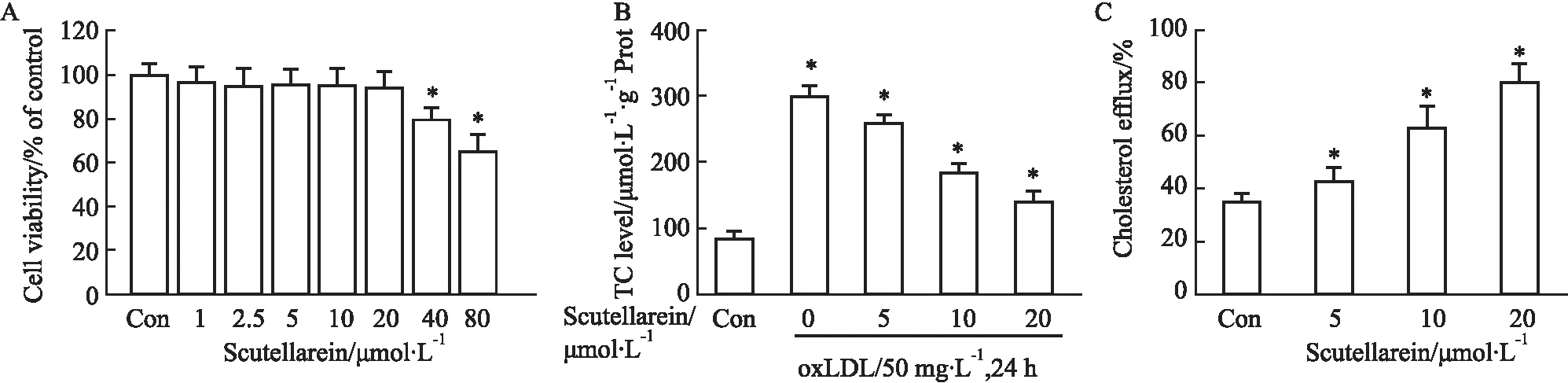
Fig 1 Effects of scutellarein on cholesterol efflux and cholesterol accumulation of macrophages n=3)
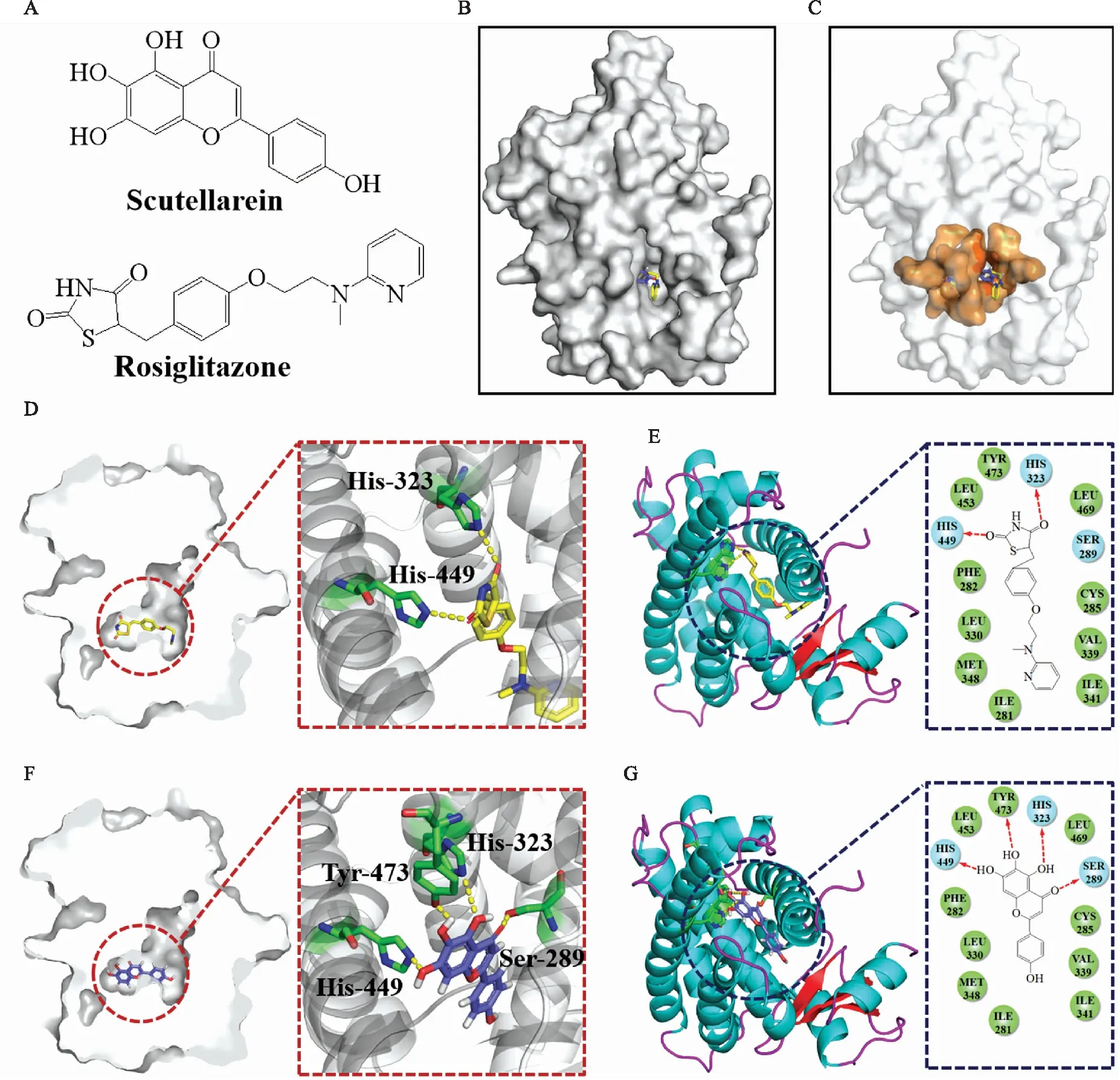
Fig 2 Molecular docking analysis of binding of PPARγ to scutellarein and rosiglitazone
2.3 野黄芩素可结合并激活PPARγ的转录活性选取罗格列酮作为阳性对照,通过ELISA方法分析比较野黄芩素及罗格列酮与PPARγ重组蛋白的结合活性。结果如Fig 3A所示,野黄芩素能够以浓度依赖的方式结合PPARγ重组蛋白。同时,在相同浓度条件下(5、10、20 μmol·L-1),野黄芩素的结合活性均明显高于罗格列酮。进一步的双荧光素酶报告基因实验结果显示(Fig 3B),野黄芩素的刺激作用可按照浓度依赖的方式激活THP-1巨噬细胞中PPARγ的转录活性,并且在相同剂量下(5、10、20 μmol·L-1),其相比于罗格列酮具有更高的激活活性。以上结果证实,野黄芩素可结合PPARγ,并激活其转录活性。

Fig 3 Effect of scutellarein on binding and transcription activity of PPARγ n=3)
2.4 野黄芩素诱导LXRα和ABCA1的蛋白表达随后,通过Western blot方法分析野黄芩素对LXRα和ABCA1蛋白表达的影响。结果如Fig 4所示,与对照组相比,野黄芩素的刺激可按照剂量依赖的方式明显提升LXRα(Fig 4A)及ABCA1(Fig 4B)的蛋白表达,此结果提示野黄芩素可通过启动LXRα-ABCA1信号通路增强巨噬细胞胆固醇外流途径。

Fig 4 Effects of scutellarein on expression levels of
2.5 PPARγ基因沉默对野黄芩素降低TC含量、增强胆固醇外流速率和LXRα和ABCA1蛋白表达的影响为了进一步明确PPARγ在野黄芩素增强巨噬细胞胆固醇外流途径中的作用,利用siRNA干扰技术,建立了PPARγ基因沉默巨噬细胞模型。如Fig 5A所示,与sramble-siRNA对照组相比,PPARγ-siRNA组细胞中,PPARγ蛋白表达水平下降约95%,表明PPARγ基因沉默细胞模型建立成功。随后,使用20 μmol·L-1野黄芩素分别刺激sramble-siRNA和PPARγ-siRNA组细胞后发现,与sramble-siRNA对照组相比,PPARγ基因沉默可明显抑制野黄芩素诱导的LXRα和ABCA1蛋白表达,并减弱了胆固醇外流途径(Fig 5B-D)。此外,在PPARγ基因沉默细胞模型中,野黄芩素对巨噬细胞胆固醇蓄积的抑制作用也被明显的逆转(Fig 5E)。以上结果证实,野黄芩素可通过激活PPARγ-LXRα-ABCA1信号通路,增强胆固醇外流途径,进而抑制巨噬细胞的胆固醇过度蓄积和泡沫化作用。
3 讨论
巨噬细胞泡沫化是AS发病过程中的核心环节,其也被认为是导致AS斑块形成以及血管狭窄的关键因素[1]。在AS早期阶段,巨噬细胞可通过细胞表面清道夫受体大量摄取oxLDL,并引发细胞内胆固醇蓄积,进而导致巨噬泡沫细胞的形成[2]。随着病情的发展,在炎症和氧化应激的作用下, caspase信号通路介导的细胞凋亡作用被启动,并导致巨噬泡沫细胞吞噬功能受损,从而使其无法清除血管内膜组织的细胞碎片。随着细胞凋亡作用的加深,巨噬泡沫细胞发生继发性坏死并不断积累,并与迁入至内膜中的平滑肌细胞共同构成了AS斑块核心,进而推动AS斑块的形成[11]。在AS晚期,AS斑块不断生长、破裂,形成血栓导致动脉血管阻塞,最终导致各种心血管疾病及并发症。胆固醇外流途径是巨噬细胞防止自身胆固醇过度积累的重要代谢途径,其可通过Apo-A1将胆固醇以高密度脂蛋白的形式转运至肝脏进行循环和再利用,以维持细胞内胆固醇的代谢平衡[2],进而避免巨噬泡沫化细胞的形成。因此,增强胆固醇外流途径对防治AS至关重要。
野黄芩素(4′, 5, 6, 7-四羟基黄酮)为野黄芩苷的苷元成分,主要分布于半枝莲、灯盏细辛、木蝴蝶及黄芩等中药材中[4]。近年来研究表明,野黄芩素具有抗炎、抗癌、抗氧化和保护神经等多种功效,并且相比于野黄芩苷,其还表现出更高的生物学利用度,因此具有较高的药用开发价值[5]。近期的报道显示,野黄芩素可通过调控血管内皮细胞的增殖以及炎症作用来改善心脑血管功能[5],但其是否能够增强胆固醇外流途径并抑制巨噬细胞泡沫化目前尚不清楚。目前,PMA诱导的THP-1巨噬细胞被广泛用于AS发病机制以及治疗方法的研究,其也是建立巨噬泡沫细胞的理想材料[12]。为了探究野黄芩素对巨噬细胞泡沫化的影响,利用50 mg·L-1oxLDL,建立了THP-1巨噬泡沫细胞模型,并发现野黄芩素可明显抑制oxLDL诱导的胆固醇过度蓄积。此外,野黄芩素的刺激作用还可加速巨噬细胞胆固醇外流途径,提示野黄芩素能够通过促进胆固醇外流途径阻止巨噬细胞的泡沫化。
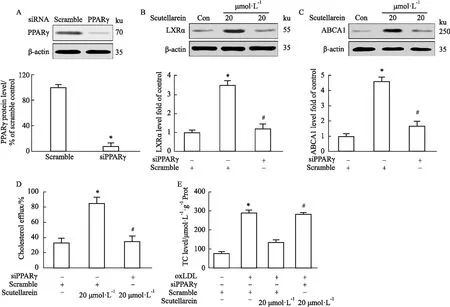
Fig 5 Effects of PPARγ knockdown on scutellarein-induced increase in cholesterol efflux and reduction in macrophage foam cell formation n=3)
过氧化体增殖物激活型受体γ(peroxisome proliferators-activated receptor γ,PPARγ)作为一种配体激活型核转录因子,在巨噬细胞胆固醇逆转运途径中发挥了关键的调控作用[13]。研究显示,PPARγ可被多种配体激活,并可与视黄醇类X受体α(retinoid X receptor α,RXRα)形成异源二聚体。该二聚体能够通过结合顺式作用元件PPRE,依次诱导下游肝X受体α(liver X receptor,LXR)及ABCA1的蛋白表达,进而加速巨噬细胞胆固醇外流途径,并抑制巨噬细胞泡沫化作用[14]。动物研究结果表明,增强巨噬细胞PPARγ的转录活性可明显减少AS斑块的形成[15]。此外,Stemerman和Brunelleschi 的研究表明,PPARγ活化后还可通过抑制核因子κB(nuclear factor kappa-B,NF-κB)的转录活性,减少炎症因子的表达,进而降低了血管的炎症反应和损伤作用[16]。由此可见,PPARγ的激活对于阻止巨噬细胞泡沫化和改善AS至关重要,其也成为治疗AS的关键靶点。目前合成类PPARγ激动剂有罗格列酮及吡格列酮等药物,但长期服用可导致患者水肿、体质量增加,甚至会产生心肌梗死的危险[17]。因此,从天然产物中寻找安全高效的PPARγ激动剂已成为当今AS研究的重点。
目前研究认为,PPARγ-LBD结构域中的T形活性口袋是PPARγ激活剂的主要结合部位[18],PPARγ激活剂与PPARγ-LBD之间的结合能越低,二者之间结合越稳定。为了进一步明确野黄芩素促进巨噬细胞胆固醇外流途径的作用机制。本文分析了野黄芩素与PPARγ的相互作用。分子对接结果显示,野黄芩素可插入至PPARγ-LBD结构域的T形活性口袋,与Ser-289、His-323、His-449及Tyr-473形成多个氢键,且相较于罗格列酮具有更低的结合自由能,提示野黄芩素能与PPARγ结合,并且结合更为稳定。ELISA、双荧光素酶报告基因以及Western blot结果进一步证实,野黄芩素能结合PPARγ,并激活其转录活性。同时,野黄芩素的刺激作用还可上调PPARγ下游LXRα和ABCA1的蛋白表达,表明野黄芩素可诱导PPARγ活化。随后,我们分析了PPARγ在野黄芩素提升巨噬细胞胆固醇外流途径中的作用。基因沉默实验结果表明,PPARγ基因沉默可明显抑制野黄芩素诱导的LXRα和ABCA1蛋白表达,并减缓了胆固醇外流途径。此外,在PPARγ基因沉默细胞中,野黄芩素对巨噬细胞泡沫化的抑制也被明显减弱。以上显示PPARγ密切参与了野黄芩素对巨噬细胞胆固醇外流途径及泡沫化形成的调控作用。
综上所述,野黄芩素可通过诱导PPARγ活化启动LXRα-ABCA1信号通路,进而增强胆固醇外流途径,最终阻止了巨噬细胞的泡沫化。本研究首次揭示了野黄芩素作为多种中药的活性成分对巨噬细胞泡沫化作用的干预效果和潜在机制,这将为今后野黄芩素用于AS的临床治疗提供理论依据。由于时间及条件的限制,本研究仅在细胞水平上分析了野黄芩素对巨噬细胞泡沫化的影响,但是其是否能够在动物水平上抑制巨噬细胞泡沫化,并阻止AS斑块形成还需要进一步验证。在今后的研究中,我们将构建AS小鼠模型,并在动物水平上验证本文所得结论。
