穗花牡荆花芽分化过程中形态和生理指标变化
2023-11-14易仁知黄清俊
易仁知,秦 俊,黄清俊*
(1 上海应用技术大学 生态技术与工程学院,上海 201418;2 上海辰山植物园城市园艺技术研发与推广中心,上海 201602)
穗花牡荆(Vitexagnus-castus),属唇形科(Lamiaceae)牡荆属(Vitex),小型落叶花灌木,原产于欧洲,主要分布于亚洲中部, 欧洲南部及地中海地区,国内多见于江浙沪一带地区栽培[1]。其蓝紫色花序大而多,花色艳丽且夏季花期长,观赏效果佳,常在湖滨、溪地、道路两侧或公园等园林配植。穗花牡荆不仅是园林应用前景好的夏花植物,且其果实药用历史悠久,已在国外广泛用于妇产科疾病治疗[2]。目前关于穗花牡荆的园林研究主要集中于育种方面[3-7],如扦插繁育、组织培养、多倍体诱导等;少数学者就栽培技术进行了阐述,仅见于遮阴、摘心、水分胁迫、多效唑处理[8-9],对其开花生理特性和花期调控技术鲜有报道。
花芽分化作为植物从营养生长向生殖生长转变的重要标志[10],划分植物花芽分化进程常分为分化初期、萼片分化期、花瓣分化期、雄蕊分化期和雌蕊分化期等多个时期,不同物种具有差异性。比如,赵通等将杏(Prunusarmeniaca)花芽分化进程划分为未分化期、分化初期、萼片分化期、花瓣分化期、雄蕊分化期、雌蕊分化期、花粉细胞期7个时期[11]。张诗艳等将海南蒲桃(Syzygiumcumini)花芽形态分化过程划分为7个阶段:花序原基分化期、小花原基分化期、萼片原基分化期、花瓣原基分化期、雄蕊原基分化期、雌蕊原基分化期和雄蕊雌蕊形成期[12]。目前,关于花芽分化的调控存在许多假说,例如碳氮比学说[13]、激素平衡[14]和信号调节假说[15]等。碳氮比学说提出碳氮比达到较高的比例才会促使花芽分化,在一理论在许多植物中得到验证[11,16]。王桢等人研究表明,较低ABA/GA3、IAA/GA3、ABA/IAA促进西红花顶芽花芽分化[17]。
据物候期观察,在上海地区穗花牡荆返青萌芽一般3月下旬开始,顶芽迅速膨大后4月初展叶,展叶后植物整体枝条、叶片加快生长,为花芽分化做准备。在江浙一带穗花牡荆一年主要有两季集中的花期,主要在6-8月份。花芽分化的数量和质量很大程度上影响着穗花牡荆的观赏品质,目前该物种花芽分化相关研究尚未报道。因此,通过划分穗花牡荆花芽分化进程和掌握不同时期形态特征,以及探究花发育过程中生理及内源激素变化,可为今后穗花牡荆的花期调控和栽培选育提供理论基础。
1 材料和方法
1.1 试验材料与处理
供试物种为上海辰山植物园所种植的5年生穗花牡荆盆栽苗,选择生长健壮,长势相似且无病虫害的植株取样,共100株。试验连续1年物候观察,从2020年12月初观察顶部冬芽的萌动情况,从冬季清晨约每隔5 d采样,春季约每隔3 d采样,直到花盛开为止。采用物候期观察和形态解剖结合观测穗花牡荆花发育过程,2021年3月24日开始返青萌芽进入营养生长期,4月初花芽逐渐进入形态分化期,4月12日开始观察到穗花牡荆花序轴原基分化,于4月中旬出现肉眼可见的分生组织,5月中旬大部分花芽现蕾。
1.2 试验方法
1.2.1 花芽形态观察
于 2021 年 3 月 24 日开始试验,连续2~3 d采样,直至现蕾。每次随机取大小一致芽体发育程度相似的20个新梢中上部顶芽,1/2数量芽体置于体视显微镜Olympus SZX16纵剖观察,确定花芽分化时期;剩余顶芽剥去叶片,固定于FAA(95%乙醇∶冰醋酸∶甲醛=9∶0.5∶0.5)溶液中,48 h后70%的酒精永久保存。固定好的花芽采用常规石蜡切片方法切片,用莱卡Leica2245切片机进行6~8 μm切片,切片后放置于莱卡H1220烘片台烘干后用甲苯胺蓝染色,置于生物显微镜Nikon BX41下拍照。在显微成像系统下将各时段解剖的花芽和石蜡切片数量进行花芽分化情况观察和数据统计,花芽阶段的划分标准为某一分化阶段花芽数量超过50%。
1.2.2 生理指标测定
根据形态解剖观察结果从花芽分化初期(2021 年 4 月6 日后)连续采样叶片和花芽用于生理指标测定,部分叶片和花芽105 ℃杀青15min,72 ℃烘干至恒重后磨样用于总碳和总氮含量测量,部分直接放入液氮罐中冷冻后转入-80 ℃超低温冰箱保存用于各项生理指标的测定。碳氮比(C/N):称取烘干后样品0.01 g,加入5 mL重铬酸钾(0.8 mol/L),加入5 mL浓硫酸油浴消煮,用0.2 mol/L的硫酸亚铁滴定,测定总碳含量。称取样品0.3 g,加入5 mL浓硫酸,消煮,分3次加入双氧水,直至消煮液呈清亮,转移至100 mL容量品,定容过滤,量取10 mL进行蒸馏,利用0.01 mol/L H2SO4进行滴定,测定总氮含量。可溶性糖、可溶性蛋白含量通过试剂盒测定,试剂盒购于索莱宝有限公司。赤霉素GA3、细胞分裂素CTK、生长素IAA以及脱落酸ABA采用酶联免疫吸附法,试剂盒及试验方法来源于上海酶联生物公司。
1.3 数据与图像处理
用Excel 2021软件进行数据整理,SPSS 25.0进行单因素分析和 Duncan’s多重比较, SigmaPlot 14.5和Prism 9软件绘制图表,图片拼接借助 Photoshop 2021软件。
2 结果与分析
2.1 穗花牡荆花芽分化过程内部形态观察
根据穗花牡荆开花物候期和花芽分化进程的观测结果,研究发现该物种在当年主要有2次集中稳定的夏季开花和秋末零星开花现象,花芽分化类型为当年多次分化型。穗花牡荆在3月展叶后迅速进行营养生长,同时受到花芽分化诱导,顶部生长锥转化为总轴花序原基;花序原基在4-6月不断进行形态分化直至花器官发育完成。据石蜡切片观察结果可划分为7个时期:未分化期、总轴花序原基分化期、初级分轴花序原基分化期、次级分轴花序原基分化期、小花原基分化期、花器官分化前期和花器官分化后期。
未分化期:2月初芽未萌动,生长点突起,混合芽形态,处于未分化期。3月中旬,芽体生长点顶端较小,基本呈圆锥型,在营养生长锥两侧的叶原基逐渐向内分化,保护内侧生长点,外层细胞排列紧密,处于花芽诱导态 (图1,A)。

A.未分化期;B.总轴花序原基分化期;C.初级分轴花序原基分化期;D.次级分轴花序原基分化期;E.小花原基分化期;F.花器官分化前期;G.花器官分化后期。Lp.生长锥;Gp.叶片;Bp.顶生花序原基;Br.初级腋生花序原基;Apm.次级腋生花序原基;1st Axm.苞片原基;2nd Axm.苞片;Fp.小花原基;Fsp.小花萼片原基;Fpp.小花花瓣原基;Pp.雌蕊原基;Sp.雄蕊原基;Se.萼片;Pe.花被;St.雄蕊;Pi.雌蕊;An.花药;Ov.子房。图A-E为整个花芽,图F-G为花芽顶端部位。
总轴花序原基分化期:叶芽生长锥呈现半圆形逐渐变宽,由花芽分化前期的花芽诱导态变为花序总轴原基分化期。靠近枝条顶端的芽体生长点有小程度的突起,肉眼很难分辨外部形态发生的变化,此时芽为混合芽。由切片可见,突起处细胞排列紧密,染色较深,原形成层由半圆形变成“八”字形,总轴原基直径为0.11 mm(图1,B)。整个生长锥趋向半圆形,长度显著增加,形成多级苞片,保护内部幼嫩的花芽分生组织。此期出现约在2021年3月14至4月9日, 4月9日观察的花芽全部进入此时期。
初级分轴花序原基分化期:于3月末腋芽表面御寒的芽鳞已基本脱落,芽体逐渐返青展叶,新梢生长出5~8个节间数,顶部和腋间混合芽直径迅速膨大。4月中旬,总轴不断伸长,花芽从混合芽中露出,肉眼可见苞片腋间有明显的分生组织,进而产生双侧分轴,为第二级花序分轴原基分化期。在总轴两侧的线形苞片腋间产生隆起的分生组织,随后发育成侧级花序,为花芽分化分轴原基。切片可见,花序分轴原基上已分化出保护幼花的苞片原基(图1,C)。此期出现盛期在4月12至4月22日。
次级分轴花序原基分化期:花梗不断增长,花蕾增大。在初级分轴花序原基两侧腋间以第一轮花序分化方式重复分化次级分轴花序原基(图1,D)。4月底花序表面包裹着多层苞片,伴随着分化逐渐伸长膨大露出十分明显的总状花序,每个总轴上末端最表层苞片不脱落,最终发育成叶片。此期出现盛期在4月25日至4月28日。
小花原基分化期:总轴花序梗和次级分轴更迅速增长,整个花序长度骤增。随后在次级分轴花序原基底部两侧按照二歧聚伞花序依次分化出小花原基(图1,E),次级分轴花序轴上着生的两侧的小花原基较中间的小花原基分化程度较晚,随着小花分化程度增加,小花数目明显增加。此期出现盛期在4月28日至5月5日。
花器官分化前期:萼片原基逐渐出现,随后覆盖花顶。花序轴上大量小花原基上两侧有小花萼片原基发生(图1,F)。此期出现盛期在5月1日到5月8日。
花器官分化后期:在与萼片相间的位置上,5枚花瓣原基同一时间分化,随后,4枚雄蕊原基同时分化。花瓣大部分内弯后,环形心皮在花顶出现,后演变为子房、花柱、柱头3部分(图1,G)。穗花牡荆花器官发生是较典型的轮状发生,萼片、花瓣、雄蕊、心皮,向心式逐轮分化,每朵小花发育顺序从花序底部向上,由中间向两侧。此期出现盛期在5月8日至5月16日。
伴随着新梢的长度增加和花序轴不断生长,小花在分轴花序上呈聚伞状花序构成次级分轴花序,且离花序顶端越远,分化越早,处同一水平位置花芽分化进程相似。花序总梗常有0~4个分轴,构成复圆锥花序。4月下旬开始分化萼片原基、花瓣原基、雄蕊原基、雌蕊原基,单个芽体开始现出两芽鳞之外和体积膨大,从外向心式发育;同时花梗不断伸长,5月16日全部现蕾,顶部花芽已经分化完成,处于育蕾阶段,即花器官逐步发育过程。所有小花依次从下向上从里往外开放。花萼钟状顶端5齿裂,花冠顶端5裂,2唇形,雄蕊4~5,柱头2裂,子房由2心皮组成。
2.2 花芽分化进程中碳氮物质含量变化
随着花芽分化时间推进,花芽和叶片可溶性糖总量整体呈现升高的趋势。由图2可知,总轴花序原基分化期前期叶片可溶性糖含量低于花芽。4月6日至4月12日,总轴花序原基分化时,叶片中可溶性糖含量第1次急剧上升,且含量高于花芽。随着腋间分生组织的出现,即初级分轴花序原基分化期,叶片中可溶性糖含量在4月15日达到最低值4 mg/g;后期次级分轴花序原基大量分化时,叶片4月25日中可溶性糖含量达到低峰值10.5 mg/g,说明花序原基分化均需消耗叶片中大量可溶性糖。4月6日至4月25日,花芽中的可溶性糖含量缓慢上升,有一定的可溶性糖积累,说明叶片内可溶性糖往花芽内运输和花芽中可溶性糖的积累有利于花芽花序原基分化。4月25日以后,叶片可溶性糖含量整体上升积累,为花器官分化做准备。4月25日至5月8日,花芽中可溶性糖含量整体上升,有利于小花原基的分化,其中萼片原基分化消耗了部分花芽内的可溶性糖含量。5月8日至5月16日,花芽中可溶性糖含量显著下降,说明花器官分化需要消耗可溶性糖。

图2 穗花牡荆花芽分化进程中可溶性糖、可溶性蛋白变化
随着花芽分化时期推进,叶片中蛋白质含量呈“降-升-降-升”变化趋势,花芽中蛋白质含量呈“升-降-升”变化趋势。总轴花序原基分化时期叶片可溶性蛋白比花芽中多,叶片中的蛋白质向花芽转移保证了花芽分化的顺利进行,花芽需要大量的蛋白质供应。4月15日至4月22日分轴花序原基叶片需要大量的蛋白质积累,为后序小花原基分化提供所需的物质和能量;5月1日至5月16日花器官分化期,说明花器官分化叶片中需要较高水平的蛋白质含量。
4月6日至4月22日,花芽中蛋白质含量持续上升,说明花序原基的分化需要大量的蛋白质积累;4月22日至5月1日,花芽内蛋白质的消耗保证了小花原基分化顺利进行;5月1日至5月16日,花芽中蛋白质含量上升而叶片中蛋白质含量下降,说明叶片的蛋白质输送到花芽促进花器官的分化。
穗花牡荆花芽分化进程中全碳、全氮含量变化如图3所示,叶片中全碳含量的变化呈升-降-稳定-升趋势,花芽中全碳含量的变化整体呈先升后降趋势。全碳含量在4月9日最低,叶片和花芽中全碳含量接近,达到平衡,大多数混合芽的生长锥逐渐从营养生长转向生殖生长,逐步上升到4月19日出现峰值。

图3 穗花牡荆花芽分化进程中全碳、全氮含量、碳氮比变化
4月19日至4月25日次级花序原基大量分化,叶片和花芽中全碳含量降低。4月25日至5月8日,花芽中碳含量迅速上升,保证小花原基大量分化。5月8日后,花芽中的全碳含量降低,叶片中全碳含量上升,与花瓣原基和雌雄蕊原基分化有关。叶片中全氮含量的变化呈先小幅度上升-下降-小幅度上升趋势,花芽中全氮含量的变化整体呈下降趋势。叶片中全氮含量在4月13日达到最高值48.75 mg/g,且在次级花序原基、小花原基以及萼片原基分化期间低于花芽中的全氮含量,5月8日后由小幅度上升。随着花芽分化推进,花芽与叶片碳氮比整体上升的趋势。5月1日花器官分化期,花芽中的碳氮比高于叶片,说明花芽中保持较高的碳氮比水平有利于花器官的分化。
综上所述,穗花牡荆花芽分化过程中的不同时期叶片和花芽中可溶性糖和可溶性蛋白质含量均有波动,总体上叶片中营养物质含量高于花芽。叶片的大量可溶性糖充足保证了花序原基分化有足够的能量来源,花芽中可溶性糖含量整体上升积累有利于花器官分化。叶片中需要较高水平的蛋白质有利于混合芽向花芽形成,花芽中大量的蛋白质积累促进了花序原基的分化,叶片的蛋白质输送到花芽保证了花器官的分化顺利进行。
2.3 花芽分化进程中内源激素含量变化及动态平衡
花芽分化进程中内源激素含量变化如图4。随着花芽分化进程的推进,叶片中IAA含量呈现波动式上升后下降,其中最低值出现在4月15日(初级分轴花序原基分化期)为85.01 μg/L,最高值出现在5月1日(小花原基分化期)为101.08 μg/L,后期IAA降低对从花器官原基大量分化有着重要的作用。ABA含量总体呈先波动时式上升后下降趋势,低含量的ABA有利于总轴花序原基形成,较高水平的ABA有利于小花原基的分化,ABA含量下降可以促进成花器官分化,其中4月15日和4月25日2个峰值分别为初级花序原基和次级花序原基分化盛期。GA3含量总体呈先上升后下降趋势,在花序分化期即将结束后处于低水平状态,说明GA3含量上升有利于花芽分化。CTK含量呈“上升-下降-上升-下降”波动状态,4月9日至4月22日,CTK含量上升有利于顶部花序原基和腋间花序原基的分化;4月22日至4月25日,CTK含量下降,此时初级花序轴上的次级花序原基大量分化。5月5日达到最高值,随后下降,这说明高水平的CTK含量有利于小花原基的分化和CTK含量下降有利于花器官原基的分化。IAA、ABA和CTK在5月1日至5月8日含量均处于较高状态,说明在这期间以上3种激素达到一定的累积,进而调控花器官分化。

图4 穗花牡荆花芽分化进程中内源激素含量变化
内源激素对穗花牡荆花芽分化的调控与不同激素动态平衡有关,如图5所示。花序原基分化期,叶片中GA3/ABA和IAA/CTK趋势变化大致相同,比值处于较低水平。ABA/CTK和ABA/IAA升高伴随着小花原基和小花萼片原基分化。花器官分化期,IAA/CTK下降。叶片中GA3/CTK、GA3/ABA和GA3/IAA的变化趋势大致相同。
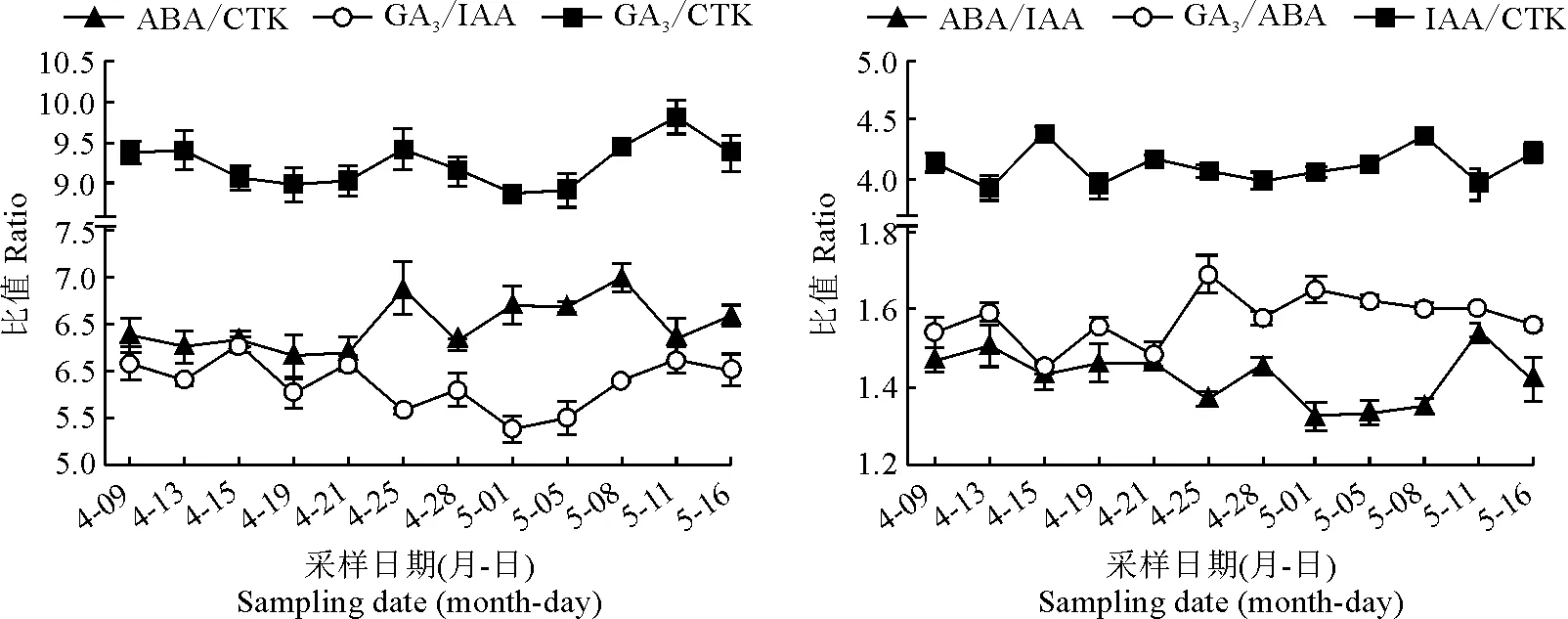
图5 穗花牡荆花芽分化进程中内源激素含量比值变化
其最低值均在5月1日,此时处于小花原基分化盛期,小花原基上的两侧花萼片原基逐渐开始分化形成花萼片,说明花萼片原基分化需要较低的GA3/CTK、GA3/ABA和GA3/IAA诱导;其最高值出现均在5月11日,此时花器官大量分化,说明花器官的分化尤其是花瓣原基、雄雌蕊原基分化需要较高的GA3/CTK、GA3/ABA和GA3/IAA;花器官的分化完成后则GA3/CTK、GA3/ABA和GA3/IAA降低,这一变化有利于花器官进一步发育。
3 讨 论
3.1 穗花牡荆花芽分化的特点
穗花牡荆作为蓝紫色系夏季时令花灌木,研究其花芽分化对花期调控和花质量提升有着重要意义。观察穗花牡荆花芽分化进程,这与复合花序的植物的研究结果[18-19]基本相符。由于穗花牡荆属于复合圆锥式的聚伞花序,且同一花序中不同的单花处于不同的分化时期和发育状态,不同空间位置花序开花时间顺序也存在差异,即枝条顶端的花序先发育成熟,花序分支底端小花和分支中间的花先开放。这与具有典型聚伞花序开花特征的皱叶膏桐花序(二歧聚伞花序)开花顺序存在差异[21]。穗花牡荆属于无限花序,不同枝位的小花原基分化期、花器官分化前期和花器官分化前期阶段存在部分的重叠现象。穗花牡荆1年有明显的两季开花现象,与皱叶膏桐[20]、麻疯树[21]、‘L6’萱草[22]的2次开花特征相似。郭娜[23]研究发现2次成花的丁香第1季的花芽属于夏秋分化型,第2季的花芽属于当年分化型;而穗花牡荆的第1季花芽属于当年分化型,其能多次诱导成花的主要原因在于花芽不需要经过低温休眠,温度和营养条件达到便可多次开花。根据花芽分化进程,可推测在分轴花序原基分化期到小花原基分化期之前,即小花原基形成之前的阶段,通过一些外源激素或物理手段影响花序与小花的数量,进行花期调控和提升花品质。
3.2 生理指标与花芽分化的关系
花芽分化是十分复杂的生理生化的过程,需要大量的碳水化合物、可溶性蛋白等营养物质积累以及碳氮物质的比例平衡来保证花芽分化顺利进行[24-25]。吴秋洁等[26]表明可溶性糖、淀粉和可溶性蛋白含量均显著上升有利于花器官发育过程。张诗艳等人[12]表明海南蒲桃花芽形态分化过程花芽可溶性糖和淀粉含量变化趋势为先降后升趋势,花芽形态分化和开花需要大量碳水化合物积累。何文广等[16]表明山鸡椒雌花在花芽分化期间从叶片部位输送大量的可溶性蛋白来供花芽分化进行。本研究的结果也表明随着穗花牡荆花芽分化时间推进,花芽可溶性糖总量整体的升高有利于花芽分化。在花芽分化前期花序原基的分化需要叶片中大量的蛋白质积累,花芽分化后期花芽中蛋白质含量上升而叶片中蛋白质含量下降说明叶片中蛋白质输送到花芽促进花器官的分化。叶片中的可溶性蛋白含量在花芽分化中呈现上升下降再上升趋势,最小值出现在花序分轴原基大量的分化时,最大值出现在花器官大量分化时。几个重要的花芽分化时刻存在峰值;其次,碳氮比学说认为,树木碳和氮的比例合适,花芽分化旺盛[11]。从碳氮比看,穗花牡荆碳氮比逐步上升,花芽中后期的碳氮比高于叶片说明花芽中保持较高的碳氮比水平有利于花器官的分化。这一点与山鸡椒的研究结果[16]一致。因此,穗花牡荆在生产上可通过调节氮肥的比例和环剥等园艺手段增加碳氮比,从而调控其花芽分化进程。
植物内源激素含量及激素平衡对花芽分化的顺利进行起着重要的调控作用。生长素对花芽分化的影响存有分歧。研究表明低浓度IAA有利于竹叶兰[26]和板栗[27]花序分化和生长。本试验IAA含量整体先上升后下降,呈波动式,低水平IAA有利于穗花牡荆的花序原基形成。GA3作为调控成花信号的重要激素之一,关于苹果、葡萄、桃、梨等植物的众多研究表明低水平GA3有利于花芽分化,高水平GA3抑制花芽分化[28],但这并不绝对。本试验穗花牡荆叶片的GA3水平在花芽分化进程持续上升累积,与山鸡椒叶片中的GA3含量趋势变化[29]一致。这可能是由于穗花牡荆植物存在大量萜类物质[30],二者在合成通路中拥有共同的前体途径[31],导致赤霉素随之累积。ABA和CTK作为促花激素,大量研究表明高水平ABA和CTK可以促进花芽分化[32-34]。从试验结果来看,低含量的ABA有利于花序原基分化,较高水平的ABA有利于小花原基的分化,ABA含量下降可以促进成雄雌蕊原基分化,与薇甘菊[35]和枣[36]花芽分化的研究相似;叶片中高水平的CTK含量有利于穗花牡荆小花原基的分化和CTK含量下降有利于其花器官原基的分化。
内源激素调控花芽分化常体现在不同种类激素的动态平衡。各类植物激素不仅会在花芽分化进程中呈现出一定的调控作用,而且还彼此拮抗或促进。多种的激素比例变化就会生成平衡状态。越来越多的学者从激素平衡角度去解释花芽分化的规律。如髙比值的ZT/IAA、ABA/IAA、ZT/GA3、ABA/GA3促进枇杷花芽分化[37];龙眼顶芽花序主轴分化前期IAA/GA3处于低水平,后期ZT/GA3升高能促进花芽分化,所有的形态分化期ZT/ABA及GA3/ABA较低[38]。在本研究中,较低水平GA3/ABA也有利于花序分化。小花原基分化期间,ABA/CTK、ABA/IAA比值升高有利于小花原基和小花萼片原基形成。GA3/CTK、GA3/ABA和GA3/IAA快速上升有利于花瓣原基、雄雌蕊原基发育。
4 结 论
通过研究花芽分化过程中的形态和生理变化,表明穗花牡荆花芽分化为一年多次分化型,其分化过程可划分为7个时期:未分化期、总轴花序原基分化期、初级分轴花序原基分化期、次级分轴花序原基分化期、小花原基分化期、花器官分化前期和花器官分化后期。相同一株植物不同枝条的花芽及相同花序中不同小花分化的进程存在差异,第1季花期后各阶段的花芽分化形态多同时出现。穗花牡荆花芽分化过程中整体叶片中碳水化合物含量比花芽高有利于提供营养,且较高的C/N比保证了花芽分化的顺利进行。花芽分化过程中,IAA、ABA、CTK和GA3整体水平上先升后降有利于花芽分化进行。低水平的GA3/ABA和IAA/CTK有利于花序的形成,ABA/CTK和ABA/IAA升高促进小花原基和小花萼片原基的分化, GA3/CTK、GA3/ABA和GA3/IAA升高促进花瓣原基、雄雌蕊原基发育。
