苹果花期至幼果期花朵和果实对低温的敏感性研究
2023-11-14杨永娥张晓煜梁小娟马梦瑶卫建国
杨永娥,张晓煜,3*,梁小娟,马梦瑶,卫建国
(1 宁夏大学 葡萄酒与园艺学院,银川 750021;2 中国气象局 旱区特色农业气象灾害监测预警与风险管理重点实验室,银川 750002;3 宁夏气象科学研究所,银川 750002)
中国苹果种植面积和产量均居世界首位,全国苹果种植区域已扩大到25个省(区、市)[1]。在苹果生产优势地区,苹果产业作为经济发展的支柱产业,为当地农业增效、农民增收做出了巨大贡献[2]。随着全球气候变暖,由春季低温引发的苹果萌芽期、花期冻害在中国苹果主栽区频繁发生。近20年来,中国苹果主产区每2~3年遭受1次轻度花期霜冻灾害,每3~5年遭受1次重度花期霜冻灾害[3]。山西吕梁吉县境内在2018年4月6日至4月7日凌晨遭受到了极其严重的倒春寒,苹果比2017年减产80%,损失5亿元以上[4]。陕西30个县域近40年苹果发生霜冻灾害的平均概率达29.72%[5]。由春季低温引发的苹果花期冻害在中国苹果生产区频繁发生,观测的数据表明,早春的霜冻和寒流之间有更强的相关性,使得苹果栽培的总体霜冻风险将保持在温暖的气候中,甚至可能增加[6]。
不同苹果品种的抗寒性存在显著差异[7-9];而10个苹果品种连续物候期花的抗寒性研究结果表明,随着物候过程的推进,花抗冻性逐渐降低,与品种和年份无关[10]。苹果作为易受春季晚霜冻害的仁果类树种之一,在苹果花期霜冻发生规律、霜冻调查以及霜冻防御方面研究较多[11-13]。宁夏地处中国西北地区东部季风边缘,对全球气温变化响应敏感。气温是影响评估产业发展、降低经济产量和效益及限制苹果栽培区域扩大的一个重要因素。据气象资料记载,宁夏地区苹果花期通常在 4 月中下旬,晚霜冻的发生时间有时会延迟至 5 月上旬,会对苹果花芽和果实造成严重冻害。花芽受冻会表现为内部鳞片褐变,花原基或部分花原基受冻害,柱头变黑干枯,严重时整个花芽干枯死亡,从而导致仅开花不结果;果实受冻表现为从果核到果肉变黑、果实脱落等现象,对宁夏苹果产业造成了巨大的危害,成为限制苹果产业发展的重要因素[14-15]。然而,关于苹果品种在花期不同阶段对低温的敏感性研究还很薄弱。离体试验因破坏了植物自身的抗逆途径,打破抗逆机制,以离体抗冻性试验取得的研究结果能否直接用于指导霜冻防御工作还颇受争议。因此,本试验以 ‘嘎啦’和‘富士’苹果品种为研究对象,以原位试验为基础,确定不同苹果品种在连续物候期(花蕾期、盛花期、坐果期和幼果期)的抗冻性变化类型,为建立精准苹果晚霜抗冻性评价方法提供参考。
1 材料和方法
1.1 试验地概况
试验于2022年4-5月苹果花蕾期、盛花期、坐果期和幼果期在宁夏银川市河东生态园艺试验中心进行,试验地位于东经106°27′41″,北纬38°22′51″,土壤类型为灌淤土,年平均气温9.5 ℃,年降水量183 mm左右,≥10 ℃活动积温3 858 ℃·d,年平均日照时间近2 942 h,全年无霜期166 d,试验期及苹果花蕾期前每月平均气温及积温情况见表1。
1.2 试验材料及仪器
1.2.1 试验材料
选用树体健壮、生长状况、园地土肥水管理、营养状况均一致且无病虫害的 12年生苹果树为研究对象,授粉方式均为人工授粉,在果树开花期用小毛刷子、橡皮头进行点授。南北行向,株行距 3.0 m×4.0 m。所选苹果品种为该区域主栽品种,分别是早熟苹果品种‘嘎啦’(砧木为新疆野苹果)和晚熟品种‘富士’(砧木为八棱海棠)。
1.2.2 仪器设备
原位试验采用宁夏回族自治区气象科学研究所研发的野外霜冻试验箱,该试验箱使用液氮作为制冷剂,箱体使用绝热泡沫材料,使用风扇完成气体对流循环。
温度控制过程中,在2个内部气体循环和外部气体循环控制下完成定时定温自然霜冻过程模拟。集温度采集、数据存储、数据显示、定时定温的自然霜冻模拟控制系统,测量精度达到±0.3 ℃,系统每秒钟采集数据1次,每分钟进行数据保存,霜箱设计图及实体图见图1。
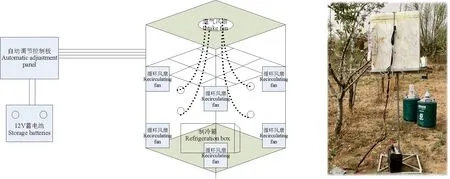
图1 野外霜冻试验箱设计图与试验图
1.3 霜冻处理
以‘嘎啦’和‘富士’为试验材料,依据文献[6]和[10]在苹果物候期花期至幼果期不同阶段均设置4个不同低温处理水平。其中,花蕾期分别为-2.0,-3.0,-4.0,-4.5 ℃;盛花期分别为-1.0,-2.0,-3.0,-4.0 ℃;坐果期分别为-1.0,-2.0,-2.5,-3.0 ℃;幼果期分别为-1.0,-1.5,-2.0,-2.5 ℃。在低温处理过程中,先在1 h内降至4.0 ℃,后以2.0 ℃/h降至设计的处理温度,在达到每个处理温度并持续1 h后,以2 ℃/h升温至环境温度,试验结束后,连续跟踪调查苹果幼果受冻率与坐果率。
1.4 测定项目及方法
1.4.1 受冻率和坐果率
受冻率是苹果花褐变数占开花总数的百分率,一般在试验结束24 h以后,统计苹果各时期供试部位褐变情况。坐果率以苹果结果数占开花总数的百分比来表示,在试验结束后,每隔7 d统计1次坐果情况,共统计3次,以最后一次为准。
1.4.2 过冷却点与结冰点
以供试样品的子房作为测定部位。取苹果花枝条置于人工霜箱(型号为SDX-20)内,将 TC-40C 型热电偶温度传感器探头固定在待测部位,设置温度曲线模拟自然界霜夜降温过程,先1 h降至4 ℃,再以2 ℃/h降至-10 ℃,采用自然升温过程升温,其原理与陈仁伟等[16]一致。
1.4.3 耐寒生理指标
用高低温交变试验箱(型号为BC1300)对供试样品进行低温冷冻处理,处理温度同1.3节。相对电导率采用电导法测定[17];可溶性糖含量采用蒽酮-硫酸比色法测定,可溶性蛋白含量采用考马斯亮蓝法测定[18]。
1.5 数据分析
采用 Microsoft Excel 2010 对数据进行预处理,Logistic 方程采用 DPS 软件进行拟合,用SPSS进行相关性和线性回归分析,Origin 2018 64Bit制图。
2 结果与分析
2.1 苹果果实不同物候期、不同降温梯度下受冻率与半致死温度变化
‘嘎啦’和‘富士’苹果花蕾期、盛花期、坐果期和幼果期各冷冻处理果实的平均受冻率调查结果如表2所示。首先,同一苹果品种同一物候期的抗寒性随着处理温度的降低而逐渐下降。例如,花蕾阶段,‘嘎啦’和‘富士’在-2.0 ℃处理下均未出现受冻情况,在-3.0 ℃处理下受冻率分别为7%和10%,在-4 ℃处理下受冻率分别为34%和47%,在-4.5 ℃时受冻率分别为58%和65%,说明2个苹果品种花蕾期受冻临界高温为-2.0 ℃。其次,同一苹果品种抗寒性随着物候过程的推进而逐渐下降。例如,在-1 ℃处理下,‘嘎啦’在花蕾阶段未表现出受冻情况,而在幼果期受冻率达到9%。另外,两品种间比较可以发现,不同苹果品种抗寒能力不同,无论处于哪种物候阶段,‘嘎啦’的抗寒性均强于‘富士’。
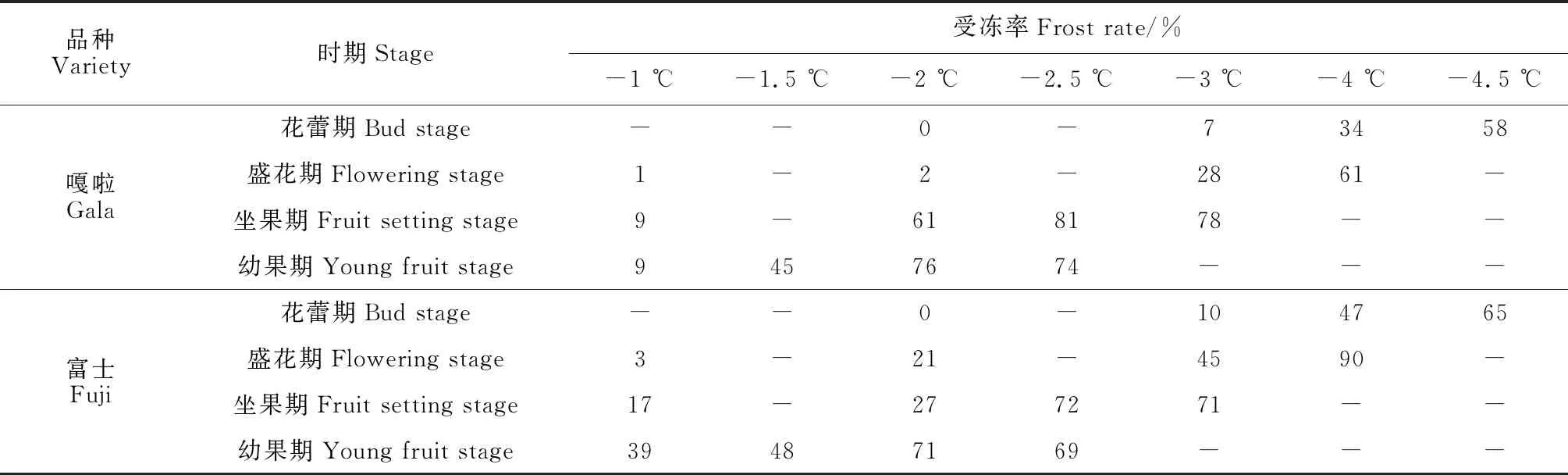
表2 苹果物候期不同阶段果实的受冻率
考虑到苹果在栽培中疏花疏果的处理,本研究将受冻率在20%以下作为自然疏花疏果过程,在20%~50%作为轻度受冻,在50%~80%作为中度受冻,在80%以上作为重度受冻。以表2中各时期的处理温度为自变量(t),以相应的受冻率为因变量(y),拟合出各品种不同时期的Logistic方程(表3),据此方程再分别计算出受冻率为20%、50%、80%时温度。
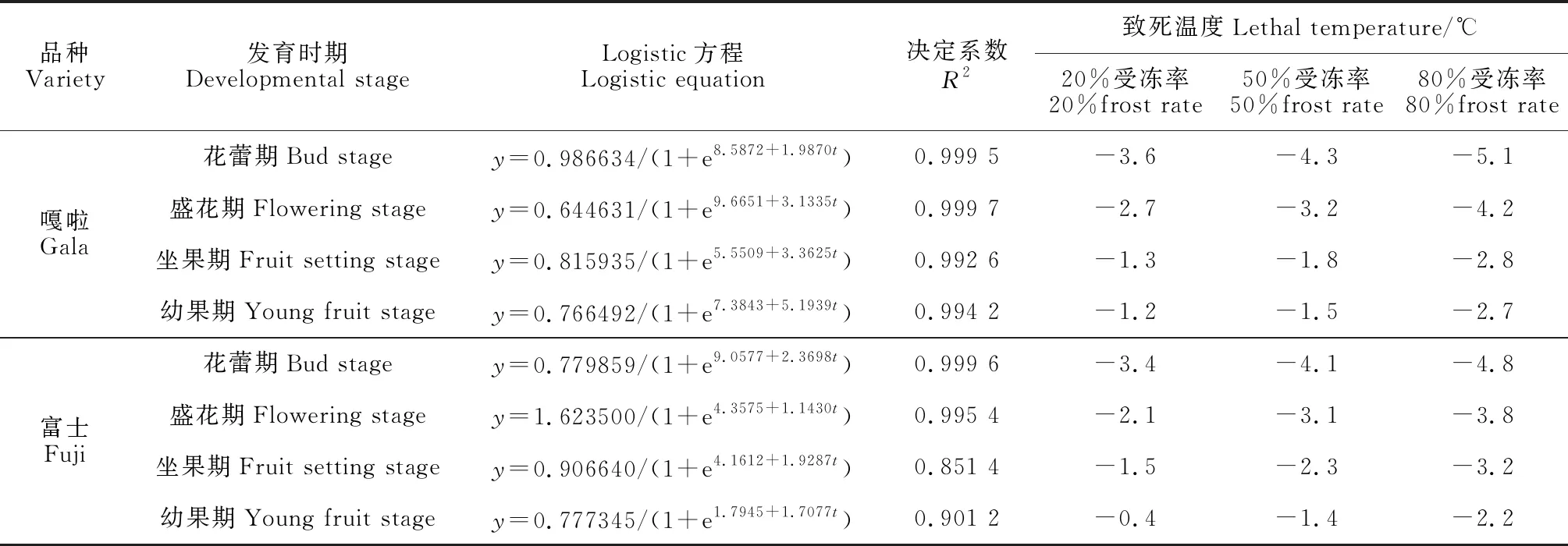
表3 苹果不同物候期受冻率与低温之间的Logistic方程及致死温度
由表3可知,苹果轻度受冻时(20%受冻率),‘嘎啦’花蕾期、盛花期、坐果期、幼果期的受冻温度分别为-3.6,-2.7,-1.3,-1.2 ℃,‘富士’则分别为-3.4,-2.1,-1.5,-0.4 ℃;苹果中度受冻时(50%受冻率),‘嘎啦’各个时期受冻温度分别为-4.3,-3.2,-1.8,-1.5 ℃,‘富士’则为-4.1,-3.1,-2.3,-1.4 ℃;苹果重度受冻时(80%受冻率),‘嘎啦’各个时期受冻温度分别为-5.1,-4.2,-2.8,-2.7 ℃,‘富士’则分别为-4.8,-3.8,-3.2,-2.2 ℃。
可见,2个苹果品种的抗寒性均随着物候期推进表现为逐渐下降的趋势,即生育期越往后越易遭受冻害;不同苹果品种抗寒性存在差异,‘嘎啦’各时期的抗寒能力始终强于‘富士’。
2.2 苹果不同物候期、不同降温梯度下抗寒性指标的变化特征
2.2.1 过冷却点和结冰点
图2显示,两苹果品种过冷却点随着物候期而逐渐上升,但在同一苹果品种不同物候期之间存在显著差异(P<0.05)。

不同小写字母表示品种及处理间差异显著(P<0.05)。
其中,‘嘎啦’过冷却点以花蕾期最低(-3.52±0.20) ℃,并与坐果期[(-3.26±0.10) ℃]、幼果期[(-3.15±0.20) ℃]存在显著差异,而与盛花期之间无显著差异;‘富士’不同物候期的过冷却点表现与‘嘎啦’相同,但其花蕾期与盛花期间差异显著。
2个苹果品种间过冷却点在花蕾期和盛花期均无显著差异,而在坐果期和幼果期差异显著(P<0.05),‘富士’坐果期和幼果期过冷却点均显著高于同期‘嘎啦’。同时,两苹果品种结冰点随着物候期先降后升,且变化幅度明显比过冷却点小,但在同一品种不同物候期间仍存在显著差异(P<0.05)。其中,‘嘎啦’坐果期结冰点[(-2.37±0.23) ℃]显著高于其它3个时期,‘富士’花蕾期和盛花期结冰点显著低于其它时期,结冰温度分别为(-2.97±0.45) ℃、(-3.03±0.16) ℃。不同品种同一物候期结冰温度均无显著差异,但‘嘎啦’结冰温度在盛花期高于‘富士’,在坐果期低于‘富士’。
2.2.2 相对电导率
从表4可知,2个苹果品种相对电导率在不同物候期均随胁迫温度的降低呈现逐渐上升的趋势。在花蕾期时,‘嘎啦’和‘富士’花器官的相对电导率在温度从室温下降至-3 ℃前均缓慢上升,但变化不显著;在温度下降至-3 ℃时分别达到37%和35%,当温度下降至-4 ℃后均迅速升高到55%~60%。在盛花期时,‘富士’花器官相对电导率在温度从室温下降至-2 ℃前缓慢上升到45%,在-2 ℃之后迅速升高至78%;而‘嘎啦’花器官相对电导率上升速度较为稳定,在-3 ℃达到50%。在坐果期时,‘嘎啦’和‘富士’花器官相对电导率在-2.5 ℃时分别达到51%和79%。

表4 不同低温胁迫下苹果相对电导率变化
在幼果时期时,在-2 ℃处理下,‘嘎啦’和‘富士’花器官相对电导率分别达到48%和80%。将苹果不同物候期花器官的相对电导率(y)对相应的胁迫温度(t)作拟合方程,从而得各物候期的Logistic方程和半致死温度(LT50),并将此半致死温度与表3中50%受冻率时的致死温度进行比较(表5)可知,‘嘎啦’、‘富士’在4个物候期50%受冻率时致死温度均高于由相对电导率拟合的半致死温度。

表5 苹果不同物候期电导率与低温的Logistic方程、半致死温度和50%受冻率温度比较
2.2.3 可溶性糖含量
可溶性糖是植物生长发育过程中所需要的主要营养物质,也是逆境胁迫下主要渗透调节物质。可溶性糖含量随低温胁迫的加剧而积累,到达一定胁迫程度后植物组织合成糖的水平逐步下降,这是植物在逆境状态下的自我保护应答机制[19-20]。
表6显示,苹果不同物候期可溶性糖含量随温度降低整体呈先上升后下降趋势。其中,在花蕾期,‘嘎啦’和‘富士’花器官可溶性糖含量在温度从室温下降至-4 ℃之前均呈现上升趋势,在-3 ℃时达到峰值(分别为28.96和21.24 mg/g),较室温下分别增加78.1%和71.0%。在盛花时期,‘嘎啦’可溶性糖含量在-2 ℃最高(24.98 mg/g),而‘富士’可溶性糖含量在-2 ℃和-3 ℃时均达到最高(19.65 mg/g)。在坐果时期,‘嘎啦’和‘富士’可溶性糖含量均在-2.5 ℃时达到最高(分别为23.64和21.11 mg/g),较室温分别增加32.2%和73.3%。在幼果期,‘嘎啦’和‘富士’可溶性糖含量在-1.5 ℃时达到峰值后,在-2.5 ℃时又呈现轻微的小范围波动上升趋势,可能是由于幼果体内其他物质小范围分解产生可溶性糖。

表6 不同低温胁迫下苹果可溶性糖含量变化
2.2.4 可溶性蛋白含量
可溶性蛋白作为植物体体内主要的渗透调节物质,在其适应逆境胁迫方面发挥着重要作用[21]。由表7知,随胁迫温度降低,两苹果品种花器官可溶性蛋白含量在花蕾期、盛花期、坐果期和幼果期整体均呈现先升高后降低的趋势。

表7 不同低温胁迫下苹果可溶性蛋白含量变化
在花蕾期,‘嘎啦’花器官可溶性蛋白含量从室温下降至-4 ℃之前整体呈上升趋势,在-3 ℃时达到峰值(4.28 mg/g),此时较室温(3.09 mg/g)增加38.5%,而‘富士’花器官可溶性蛋白含量在-4 ℃达到峰值(4.31 mg/g),较室温增加20.7%。
在盛花期,‘嘎啦’和‘富士’花器官可溶性蛋白含量均在-3 ℃达到峰值,分别为4.11,4.09 mg/g。在坐果期, ‘嘎啦’可溶性蛋白含量在-2.5 ℃时达到峰值(2.78 mg/g),‘富士’可溶性蛋白含量-2 ℃时达到峰值(2.34 mg/g);‘嘎啦’和‘富士’可溶性蛋白含量在-3 ℃时与室温下基本一致。在幼果时期,‘嘎啦’可溶性蛋白含量呈现升高-降低-升高的趋势,在-1 ℃最高(2.86 mg/g),较室温下增加14.3%,而‘富士’可溶性蛋白含量在-1.5 ℃达到最高,较室温下仅增加4.7%。同时,随着物候期的后延,两品种可溶性蛋白含量呈现下降的趋势,后2个时期明显低于前2个时期,并且不同苹果品种应对逆境的反应程度不同,‘富士’变化幅度更大。
2.3 苹果物候期致死温度及各抗寒指标之间的相关性
2个苹果品种物候期致死温度及各抗寒指标之间的相关分析结果(表8)显示,花器官半致死温度与受冻率呈极显著正相关(r= 0.909**),与过冷却点温度、结冰点温度呈显著正相关(r= 0.746*,0.796*);受冻率与过冷却点温度、结冰点温度均呈显著正相关(r= 0.741*,0.715*),与可溶性蛋白含量呈显著负相关(r= -0.874*);过冷却点温度、结冰点温度均与可溶性蛋白含量显著负相关(r= -0.896*,-0.939*);而可溶性糖含量与其余指标均无显著性相关。
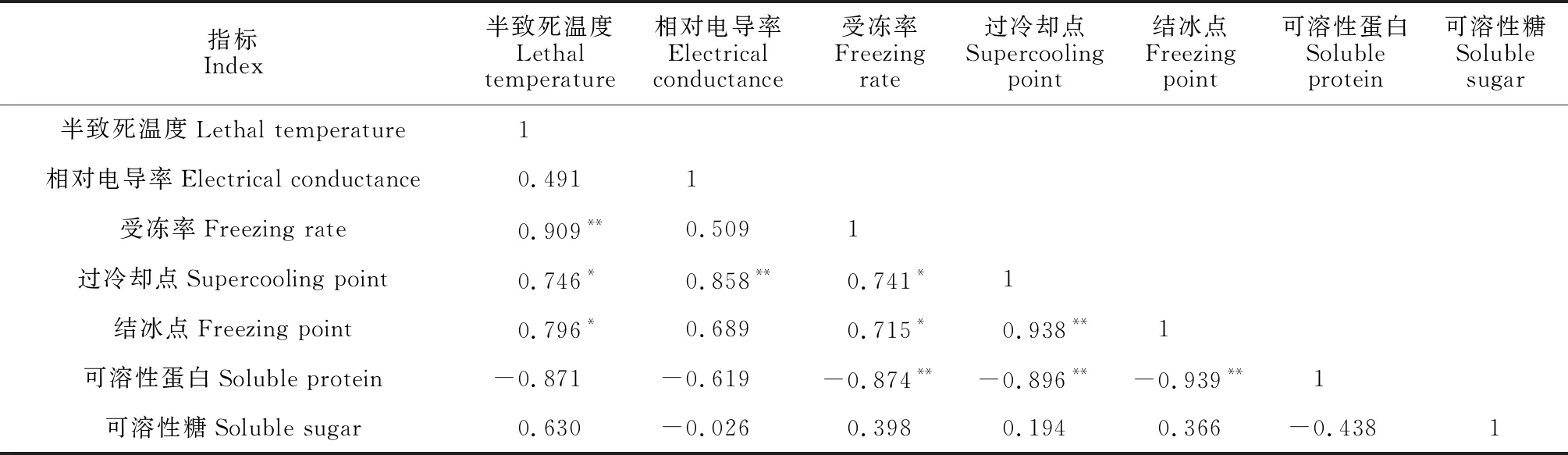
表8 抗寒性指标相关系数矩阵
以上结果说明半致死温度、过冷却点和结冰点温度越高,受冻率越高,抗寒性越差;而可溶性蛋白含量越高,供试部位受冻程度越低。
2.4 苹果不同物候期抗寒能力判断指标解析
采用多元线性回归分析法,将2个苹果品种4个时期子房受冻率作为因变量(y),将同时期相对电导率(x1)、过冷却点(x2)、结冰点(x3)、可溶性蛋白(x4)和可溶性糖(x5)作为自变量,构建多元线性回归模型(表9)。结果表明,调整后R2=0.714,说明模型解释度较好;方差分析中,F=16.480,P=0.000<0.01,说明此模型具有统计学意义。同时,从不同自变量的标准化系数及显著性P值发现,本研究涉及的4个物候期的相对电导率和可溶性蛋白含量是造成子房受冻率差异的主要因素,其次为结冰点温度,而过冷点温度和可溶性糖含量的影响相对较小。以上研究结果表明,衡量苹果花期、果期抗寒性的最主要指标是子房的相对电导率和可溶性蛋白含量,但结冰点温度、过冷点温度、可溶性糖含量等生长发育指标也是衡量苹果花蕾期至幼果期抗寒性的重要指标。

表9 多元线性回归模型
3 讨 论
本研究以原位试验为基础,运用野外霜冻试验箱对2个苹果品种花期至幼果期做降温处理,模拟自然界霜冻过程,在试验处理1周后统计各物候期子房受冻率,从而更精准地反映不同物候期抗寒能力。结果表明:不同苹果品种抗寒能力不同,‘嘎啦’抗寒能力强于‘富士’。 李晓龙等[22]研究不同品种苹果花期抗寒能力结果表明,‘嘎啦’过冷却点温度较低,其开花持续时间、花量及全开花比率与其他红色品种相比并无差异,说明其单个花朵自身的抗寒性较强,树体花期抗寒能力也较强。
本研究以受冻率建立 Logistic 方程,统计2个苹果品种轻度、中度、重度受冻临界温度,结果表明:随着物候期的推进,2个苹果品种轻、中、重度受冻临界温度呈现升高的趋势。Proebsting等[23]研究了‘红美味’苹果生殖器官的抗冻性,并确定了 LT50值,发现随着物候期推移,其耐寒性逐渐恶化,有轻微波动;红花期 LT50值为-4.4 ℃,球囊期为-3.4 ℃,开花期为-3.3 ℃,盛花期为-3.7 ℃,末花期为-2.7 ℃。本研也发现,同一苹果品种不同物候期抗寒能力不同,并表现为花蕾期>盛花期>坐果期>幼果期。有学者研究不同苹果品种在花期不同阶段的晚春抗寒性也表明,随着物候期推进,苹果抗寒能力表现出逐渐下降的趋势[24]。在苹果开花至幼果时期,花蕾期最耐寒,在开花时只能忍受零下几度的低温,因为此时雌蕊和胚珠是最敏感的部分,其他器官也会不同程度受损[25]。
相对电导率作为衡量植物抗寒性的重要指标,研究表明,果树根系、花朵、枝条等的抗寒性与电导率密切相关[26-28]。本研究中,抗寒性强的‘嘎啦’花器官的相对电导率值在各时期、不同低温处理下均低于‘富士’,其变化范围为 23.32%~70.21%,且变化趋势较为平缓,而抗寒性弱的‘富士’相对电导率值较高,并且在(-2)~(-4) ℃处理时急剧上升,说明其此时细胞膜便开始严重受损,稳定性变差。杨豫等[29]利用电导率建立的Logistic 方程和半致死温度可快速评价不同葡萄品种枝条抗寒性。本研究测定了苹果花期至幼果期在不同温度下的相对电导率,探究其对逆境的适应性和抵抗能力。结果显示,在逆境初期,花器官相对电导率呈缓慢上升趋势,随着温度降低达到一定值后显著增加,随后趋于稳定,呈现“S”形曲线。低温处理结果显示,苹果花期至幼果期对轻度逆境具有一定的适应过程,温度下降至-4 ℃时,苹果花期至幼果期相对电导率均超过50%,说明苹果花器官和幼果的膜系统在低温胁迫下遭受了一定的伤害,而且苹果花器官和幼果的伤害程度表现出显著的差异,后期相对电导率增加缓慢,证明在重度逆境条件下细胞膜已经完全破损,电解质完全外流。根据低温下等温差处理运用Logistic 方程拟合,结果得出‘嘎啦’花蕾期、盛花期、坐果期、幼果期半致死温度(LT50)分别为-4.6 ℃,-4.1 ℃,-2.8 ℃,-2.6 ℃,‘富士’各时期半致死温度分别为-4.2 ℃,-2.9 ℃,-2.3 ℃,-1.8 ℃,进一步印证了随着物候期推移,苹果供试部位抗寒能力逐渐变弱。
随着过冷却点温度的降低,植物能够在较低的温度下快速结冰,说明植株抗寒能力越强[30]。本研究发现,‘嘎啦’苹果花蕾期至幼果期过冷却点温度分别为-3.6,-3.5,-3.3,-3.2 ℃, 而‘富士’过冷却点温度分别为-3.6,-3.3,-2.8,-2.6 ℃,一旦温度降至过冷却点,苹果供试部位开始释放热量,导致温度急剧上升达到峰值,此时的峰值为结冰点。值得注意的是,‘嘎啦’苹果各个时期过冷却点温度均低于‘富士’,进一步证明了‘嘎啦’苹果相较于‘富士’具有更强的抗寒能力。此外,随着物候期推进,‘嘎啦’苹果过冷却点温度呈现升高的趋势,这也表明果树的抗寒性在物候期推进的过程中逐渐降低。
本研究发现,2 个苹果品种、4 个物候期花器官可溶性糖含量随低温处理均呈先上升后下降趋势,这与李呈呈等[31]对6个彩叶桂品种对低温胁迫的生理响应、Yan等[32]对苜蓿抗寒性生理特性的研究中可溶性糖含量变化结果一致。不同苹果品种,在同一时期同一低温处理下供试部位可溶性糖含量达到最大值的低温不同,这可能与品种间特性不同有关。在达到一定低温时,可溶性糖含量与可溶性蛋白呈下降趋势,可能是因为苹果花器官或幼果遭受严重冻害胁迫后,细胞内保护酶系统等遭到严重破坏。另外,本研究对苹果各物候期霜冻指标相关性与多元线性回归表明,半致死温度和可溶性蛋白含量与抗寒性高度相关。研究表明,结缕草[31]和枣树[32]的可溶性蛋白含量与其抗寒性呈极显著相关。本研究中‘嘎啦’各时期、不同温度处理下可溶性蛋白含量均低于‘富士’,说明‘富士’的可溶性蛋白调节机制比‘嘎啦’对低温更敏感,两品种都以通过提高可溶性蛋白含量来抵御低温胁迫。曹建东在葡萄砧木抗寒性生理指标比较中也发现这一现象[33]。
4 结 论
2个苹果品种开花物候期不同阶段对低温的敏感性鉴定结果表明:
(1)‘嘎啦’花蕾期、盛花期、坐果期、幼果期重度受冻温度分别为-5.1,-4.2,-2.8,-2.7 ℃,‘富士’分别为-4.8,-3.8,-3.2,-2.2 ℃。
(2)不同苹果品种抗寒能力不同,‘嘎啦’抗寒性强于‘富士’。
(3)同一苹果品种随着物候期推进,抗寒性逐渐下降,抗寒能力表现为花蕾期>盛花期>坐果期>幼果期。
(4)传统抗寒生理指标中,半致死温度和可溶性蛋白更适用于表征受冻状况,其次为过冷却点和结冰点温度等抗寒性指标。
本研究仅从生理指标方面对苹果品种抗寒能力进行评价,而未考虑果树抗寒机理研究,为了进一步证实以上观点,今后将开展基于冷胁迫下不同苹果品种、不同物候期各器官的转录组差异性研究,从分子角度探索抗寒机制,以期为苹果栽培生产提供更科学指导。
