β-谷甾醇对小鼠乳腺上皮细胞乳蛋白和乳脂肪合成的影响
2023-11-01刘莉莉
■ 刘莉莉 陈 敏
(黑龙江中医药大学药学院,黑龙江哈尔滨 150040)
哺乳动物乳汁是最适合新生动物健康生长发育的天然营养物,乳汁含有丰富的蛋白质和脂肪,这些乳营养成分的合成与乳腺上皮细胞一些重要功能基因及信号转导通路的调控密切相关。乳蛋白主要由酪蛋白和乳清蛋白组成,其中酪蛋白约占乳蛋白的80%,故其含量的高低常作为评价乳汁营养价值的重要指标。雷帕霉素靶点(mTOR)信号途径以及酪氨酸激酶2/信号转导和转录激活因子5(JAK2/STAT5)信号途径主要调控乳蛋白的合成,而且mTOR 信号通路与JAK2/STAT5信号通路可相互影响协同调控乳蛋白的合成[1]。乳脂的主要成分是三酰甘油,主要由脂肪酸和磷酸甘油在乳腺上皮细胞中合成。过氧化物酶体增殖物激活受体(PPARG)和固醇调节元件结合转录因子1(SREBF1)是乳脂肪合成与分泌过程中最重要的两个调控因子,在乳脂肪合成基因网络中处于枢纽位置,二者可通过结合靶基因,影响乳腺对脂肪酸的摄取、转运、从头合成和酯化过程[2]。
提高乳质量及泌乳量是目前哺乳动物亟待解决的重要问题,天然药用植物及其提取物具有低毒、无残留、不易产生耐药性等优点,用于增乳有其独特优势。研究发现低剂量葛根素能增强小鼠乳腺乳蛋白和乳脂肪合成相关蛋白表达,提升乳品质[3]。槲皮素可以改善乳质量,提高小鼠乳蛋白、乳糖和乳脂肪合成相关基因的表达[4]。β-谷甾醇作为一种植物甾醇,是很多药用植物的重要组成成分。临床试验研究证实β-谷甾醇具有降胆固醇、降血糖、抗氧化、抗炎、抑菌和抗癌等多种生物活性。此外,研究发现β-谷甾醇还可通过雌激素受体途径调节下游靶基因的表达而发挥植物雌激素样活性的作用[5-6]。植物雌激素是许多植物中的天然化学成分,结构类似于雌激素17-β-雌二醇,可以与动物雌激素受体结合,发挥雌激素样活性[7]。许多增乳药用植物的活性成分中含有植物雌激素,其可通过调节泌乳相关激素的活性,促进乳腺发育,进而影响动物泌乳性能。然而关于β-谷甾醇对小鼠乳腺上皮细胞(HC11)乳蛋白、乳脂肪的合成是否有影响尚未见报道。
本研究以HC11 细胞为模型,采用MTT 法检测不同浓度β-谷甾醇对HC11细胞增殖能力的影响,qRTPCR 技术检测β-谷甾醇作用的HC11 细胞中乳蛋白和乳脂肪合成相关基因的表达情况,为探讨β-谷甾醇对HC11细胞乳蛋白和乳脂肪合成的调节机理提供理论基础,为利用天然植物活性成分提高哺乳动物乳品质提供参考。
1 材料与方法
1.1 材料
小鼠乳腺上皮细胞(HC11)购自北京北纳创联生物技术研究院。
β-谷甾醇购自成都瑞芬思生物科技有限公司;MTT 法细胞活力检测试剂盒购自上海碧云天生物技术有限公司;RPMI-1640 干粉、胎牛血清(FBS)购自Gibco 公司;Trizol 购自Invitrogen 公司;Prime ScriptTMRT reagent Kit 试剂盒、SYBR Premix Ex TaqTM试剂盒购自TaKaRa公司。
1.2 HC11细胞的培养
37 ℃、5% CO2培养条件下,利用生长培养基(RPMI-1640+10% FBS)培养HC11 细胞,每隔24 h 更换新鲜培养基。
1.3 β-谷甾醇对HC11细胞活力影响的检测
试验分为对照组(接种HC11细胞但不加药处理)和不同浓度(5、10、20、40、80、100、120 μmol/L)β-谷甾醇组。每组5 个重复孔。于96 孔板中接种对数生长期的HC11 细胞(1×104个/孔),培养过夜,更换新鲜培养基并添加β-谷甾醇,作用24 h 后,各孔加入10 μL MTT(5 mg/mL)继续培养4 h,吸出培养液,各孔分别加入100 μL DMSO,振荡10 min,酶标仪检测490 nm 的各孔吸光度(OD)值并进行计算,试验重复3次。
1.4 β-谷甾醇对HC11 细胞乳蛋白和乳脂肪合成相关基因表达影响的检测
取对数生长期的HC11 细胞接种于6 孔板中,待培养板中细胞融合度达到70%~80%时,更换新鲜培养基,对照组HC11细胞添加正常细胞培养基,各试验组HC11 细胞在添加正常细胞培养基的基础上,同时分别添加不同浓度的β-谷甾醇(5、10、20 μmol/L),每组设置5 个重复孔。β-谷甾醇作用细胞48 h,收集细胞,Trizol 法提取RNA,根据Prime ScriptTMRT reagent Kit 试剂盒说明书进行反转录,利用qRT-PCR 试剂盒(SYBR Premix Ex TaqTM)检测HC11 细胞CSN1S1、CSN2、CSN3、mTOR、STAT5、JAK2、S6K1、4EBP1、SREBF1、PPARG、PPARGC1A、ACC、FAS、SCD基因的mRNA 表达水平。各基因的引物序列见表1,β-actin为内参基因。采用2-ΔΔCt相对定量的方法计算各基因的mRNA表达丰度。

表1 实时荧光定量 PCR引物序列
1.5 数据处理
试验数据均以“平均数±标准差”表示;采用SPSS 22.0 软件对试验数据进行单因素方差分析,P<0.05 表示具有统计学差异。
2 结果与分析
2.1 β-谷甾醇对HC11细胞活力的影响
不同浓度β-谷甾醇对HC11 细胞活力的影响如图1 所示。与对照组相比,20 μmol/L 以内的β-谷甾醇对HC11 细胞活力均无显著影响(P>0.05),而β-谷甾醇添加量达到40 μmol/L 后,细胞活力随β-谷甾醇添加浓度的升高而显著下降(P<0.05)。后续试验采用对HC11 细胞活力无抑制效果的5、10、20 μmol/Lβ-谷甾醇处理细胞,以探究其对HC11 细胞乳蛋白和乳脂肪合成相关基因表达的影响。

图1 β-谷甾醇对HC11细胞活力的影响
2.2 β-谷甾醇对HC11 细胞酪蛋白及乳蛋白合成相关基因表达的影响
不同浓度β-谷甾醇对HC11 细胞酪蛋白基因mRNA 表达的影响如图2 所示,与对照组相比,5、10、20 μmol/L 的β-谷甾醇均能显著提高HC11 细胞CSN1S1、CSN2、CSN3基因mRNA 表达水平(P<0.05),其中5 μmol/Lβ-谷甾醇组细胞CSN1S1、CSN2、CSN3基因mRNA表达水平相对更高。

图2 β-谷甾醇对HC11细胞酪蛋白基因mRNA表达的影响
不同浓度β-谷甾醇作用下HC11 细胞中乳蛋白合成信号通路(JAK2/STAT5 和mTOR)相关基因mRNA表达水平的变化如图3所示,与对照组相比,5、10、20 μmol/L 的β-谷甾醇组HC11细胞STAT5、JAK2、mTOR和S6K1基因mRNA 表达水平均显著升高(P<0.05)。其中5 μmol/Lβ-谷甾醇组细胞STAT5和JAK2基因mRNA 表达水平相对最高,10 μmol/Lβ-谷甾醇组细胞mTOR基因和S6K1基因mRNA 表达水平更高。与对照组相比,5 μmol/L 和10 μmol/L 的β-谷甾醇组HC11细胞4EBP1基因mRNA表达水平无显著变化(P>0.05),而20 μmol/L 的β-谷甾醇能显著提高HC11细胞4EBP1基因的mRNA表达(P<0.05)。

图3 β-谷甾醇对HC11细胞乳蛋白合成信号通路相关基因mRNA表达的影响
2.3 β-谷甾醇对HC11 细胞乳脂肪合成相关基因表达的影响
不同浓度β-谷甾醇对HC11 细胞脂肪酸合成相关基因mRNA水平影响的检测结果如图4所示,与对照组相比,5、10、20 μmol/L 的β-谷甾醇均能明显上调HC11细胞ACC基因和SCD基因的mRNA表达水平(P<0.05),其中5 μmol/Lβ-谷甾醇组细胞ACC和SCD基因mRNA表达水平相对更高。但与对照组相比,5 μmol/L和10 μmol/L 的β-谷甾醇对HC11 细胞FAS基因表达无显著影响(P>0.05);而20 μmol/L 的β-谷甾醇能显著提高FAS基因mRNA的表达(P<0.05)。
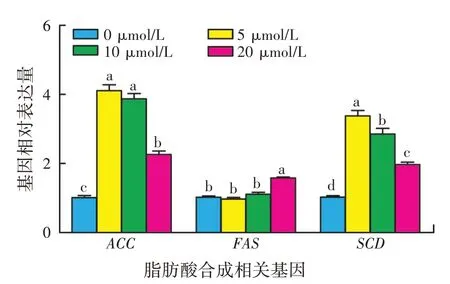
图4 β-谷甾醇对HC11细胞脂肪酸合成相关基因mRNA表达的影响
不同浓度β-谷甾醇作用下HC11 细胞中乳脂肪合成关键调控因子基因(SREBF1和PPARG)及其辅助因子PPARGC1A基因mRNA 表达水平的变化如图5所示,与对照组HC11 细胞相比,5、10、20 μmol/L 的β-谷甾醇均能显著提高HC11 细胞SREBF1、PPARG和PPARGC1A基因mRNA 表达水平(P<0.05)。其中5 μmol/Lβ-谷甾醇组细胞PPARG基因和PPARGC1A基因mRNA 表达水平相对最高,10 μmol/Lβ-谷甾醇组细胞SREBF1基因mRNA表达水平相对最高。
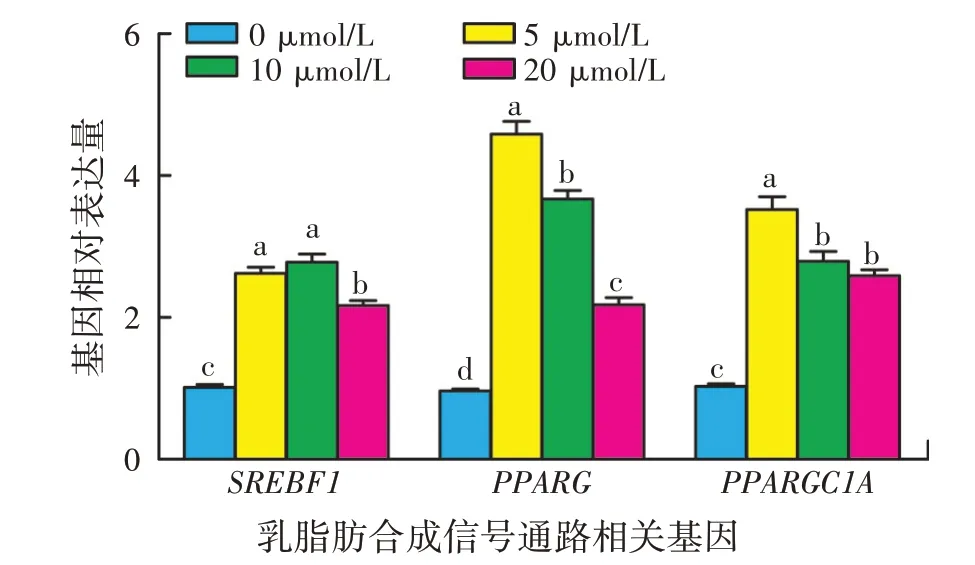
图5 β-谷甾醇对HC11细胞乳脂肪合成信号通路相关基因mRNA表达的影响
3 讨论
乳腺上皮细胞是乳蛋白、乳脂肪等营养物质合成的主要场所。哺乳动物的泌乳功能与乳腺上皮细胞的数目以及其分泌与合成乳成分的能力密切相关。郭惠中[8]研究表明低剂量β-谷甾醇对瘢痕疙瘩成纤维细胞活性无显著影响,而高剂量β-谷甾醇表现出明显抑制作用。王凯等[9]研究发现随着β-谷甾醇添加剂量的升高,肝癌细胞Hep3B和HepG2的细胞活力逐渐降低。与前人的研究结果一致,本研究发现低浓度的β-谷甾醇对HC11 细胞活力无明显影响,而高浓度β-谷甾醇对HC11 细胞活力具有显著抑制作用,因此后续试验选择对HC11 细胞活力无抑制作用的β-谷甾醇剂量来研究其对HC11细胞泌乳功能的影响。
乳蛋白主要由酪蛋白和乳清蛋白组成[10]。乳中含量最高的蛋白质是酪蛋白,约占总乳蛋白的80%,主要分为4种:αs1-酪蛋白(CSN1S1)、αs2-酪蛋白(CSN1S2)、β-酪蛋白(CSN2)以及κ-酪蛋白(CSN1S3)[11]。乳蛋白合成主要由2 条信号通路调控,分别是在催乳素或其他泌乳相关细胞因子作用下从基因转录水平调节乳蛋白合成的JAK2/STAT5信号通路和从蛋白质翻译水平调节乳蛋白合成的mTOR 信号通路。当催乳素等配体与乳腺细胞表面的受体结合后激活JAK2,活化的JAK2 磷酸化转录因子STAT5,激活的STAT5 蛋白形成二聚体,进入细胞核调控下游乳蛋白相关基因的表达,促进泌乳以及乳蛋白的合成[12]。激活的mTOR磷酸化其下游效应因子真核细胞翻译启动因子4E 结合蛋白1(4EBP1)和核糖体蛋白S6 激酶1(S6K1),被激活的S6K1 能增强含嘧啶基因mRNA 的翻译效率,调节乳蛋白的合成[13]。而真核细胞翻译起始因子4E(eIF4E)与未磷酸化的4EBP1 结合,对蛋白质的翻译产生抑制作用,4EBP1 经mTOR 磷酸化后会与eIF4E脱离,减弱4EBP1 和eIF4E 结合对蛋白质翻译的抑制作用,促进乳蛋白的合成[14]。研究发现具有植物雌激素作用的芦丁能显著提高奶牛乳腺上皮细胞CSN1S1基因和CSN2基因的mRNA 表达,对乳蛋白合成具有促进作用[15]。作为黄酮类植物激素的槲皮素能显著上调产后缺乳小鼠CSN2基因的表达量[4]。植物雌激素葛根素可能通过催乳素受体激活JAK2/STAT5a 信号通路提高β-酪蛋白的表达,实现对小鼠乳腺泌乳过程的调控[3]。和雌激素、催乳素作用相似的王不留行能提高磷酸化STAT5、mTOR及S6K1的表达,进而促进奶牛乳腺上皮细胞乳蛋白的合成[16]。与以往的研究结果类似,本试验结果表明,添加低剂量β-谷甾醇能显著提高CSN1S1、CSN2和CSN3的mRNA 水平,同时显著上调JAK2、STAT5、mTOR和S6K1的mRNA 表达,但低剂量β-谷甾醇对4EBP1的mRNA 表达无显著影响。这些结果提示低剂量的β-谷甾醇可能发挥植物雌激素的作用,激活HC11细胞JAK2/STAT5通路和mTOR 通路,进而促进乳蛋白的合成。但相比较而言,高剂量的β-谷甾醇可能会减弱乳腺上皮细胞合成分泌乳蛋白。
SREBF1 是调节乳脂肪合成的关键转录因子,参与调控乳脂肪合成过程中多种转运蛋白和关键酶的表达,影响乳腺三酰甘油的合成和分泌[17]。Ma等[18]研究发现SREBF1基因的低表达会抑制脂肪酸从头合成相关基因ACC、FAS、SCD和脂肪酸结合蛋白FABP3的表达 。PPARG属于核激素受体家族中的配体激活受体,可通过调节乳腺中与脂肪酸摄取、活化、运输、从头合成、去饱和、三酰甘油合成相关的生脂基因表达及SREBP1的表达而在乳脂肪合成中发挥关键作用[19]。PPARGC1A是PPARG基因的共激活因子,它通过与转录因子PPARG 的互作而结合在靶基因的启动子区,协助调节基因表达[20]。Kadegowda 等[21]使用PPARG 激动剂罗格列酮处理奶牛乳腺上皮细胞,发现脂肪酸从头合成基因(ACC、FAS、SCD)及转录调控基因(SREBF1)的表达量均上调。乳脂肪合成的过程也受到mTOR信号通路的调节,激活的mTOR 信号通路可以通过eIF4E、4EBP1 和S6K 等下游信号分子,上调SREBF1 和PPARG 等转录调控因子的表达以促进乳脂肪合成[22-23]。研究发现类激素物质大豆异黄酮能明显促进奶牛乳腺上皮细胞β-酪蛋白和三酰甘油的合成和分泌[24]。槲皮素可以提高产后缺乳小鼠乳腺脂肪酸合成相关基因FAS和SCD的表达[4]。王不留行黄酮苷能上调mTOR和SREBF1c 的表达,促进奶牛乳腺上皮细胞β-酪蛋白和三酰甘油合成[25]。与以前的研究结果相似,本研究发现β-谷甾醇上调乳脂肪合成转录因子SREBF1、PPARG及辅助因子PPARGC1A基因表达的同时,也促进了ACC、FAS、SCD基因的表达,提示β-谷甾醇可能通过激活mTOR 信号通路调控以SREBF1与PPARG 为核心的乳脂肪合成基因网络,上调脂肪酸合成关键酶相关基因的表达,进而促进乳腺三酰甘油的合成与分泌。
4 结论
β-谷甾醇可通过JAK2/STAT5 和mTOR 信号通路调节酪蛋白基因CSN1S1、CSN2和CSN3的表达;也可上调PPARG 与SREBF1 乳脂肪合成关键转录因子的表达进而调控脂肪酸合成相关基因ACC、FAS和SCD的表达。当β-谷甾醇添加浓度为5~10 μmol/L 时,对小鼠乳腺上皮细胞乳蛋白和乳脂肪合成相关基因表达的促进效果较好。
