乳酸对非小细胞肺癌细胞线粒体自噬的影响※
2023-10-12王迦乐黄登亮张耀刚
王迦乐,黄登亮,张耀刚,姜 军
(1.青海大学附属医院中心实验室,西宁 810001;2.青海大学研究生院,西宁 810016;3.青海大学附属医院肿瘤科,西宁 810001)
乳酸是肿瘤重要的代谢产物,低氧又是肿瘤的普遍特征,两者之间的相互作用是否会对肿瘤细胞线粒体功能产生影响?本研究通过相关研究初步探讨乳酸对人非小细胞肺癌细胞线粒体自噬的影响。
1 材料与方法
1.1 材料
1.1.1 细胞株
人正常肺上皮细胞(BEAS-2B)、人非小细胞肺癌细胞(A549)购自中国科学院细胞库。
1.1.2 试剂与仪器
乳酸(Sigma-Aldrich公司,L6402型),乳酸检测试剂盒(Solarbio公司,A019-2型),二甲基亚砜DMSO(Solarbio公司,D8370型),胎牛血清(Biological Industries公司,04-001-1ACS型),0.1%胰酶(Gibco公司,27250018型),1640培养基(Gibco公司,23400-021型),FastStart Universal SYBR GreenMaster(Roche公司,04913914001型),MitoTracker®Deep Red FM线粒体深红色荧光探针(Thermor Frsher Scientific公司,M2247型),CytoPainter LysoView Blue溶酶体染料(abcam公司,ab176825型),LC3B抗体(abcam公司,ab192890型),β-Actin 抗体(boster公司,BMO627型),TOM20抗体(abcam公司,ab56783型),HSP60抗体(Cell Signaling Technology公司,12165S型)。细胞成像微孔板检测系统(BioTek公司,Cytation5型),实时荧光定量PCR仪(ROCHE公司,ROCHE Light Cycler 480Ⅱ型),超微量核酸蛋白浓度测定仪(Nanodrop公司,Nanodrop 2000C型)。
1.2 方法
1.2.1 细胞培养与分组
将BEAS-2B、A549细胞培养于含10%胎牛血清的1640培养基中,置细胞培养箱(5%CO2,37℃,95%湿度)培养,2~3 d更换培养基,细胞汇合度达到80%左右时用0.1%胰酶消化传代,取对数生长期的细胞接种于12孔板或6孔板。将BEAS-2B、A549细胞分为常氧组(21%O2,5%CO2,37℃)、低氧组(1%O2,5%CO2,37℃)。将乳酸处理过的BEAS-2B、A549细胞分为常氧组(Lac-0、5、10 mM)、低氧组(Lac-0、5、10 mM)。
1.2.2 乳酸测定
将BEAS-2B、A549细胞接种于12孔板中,每孔细胞数为5×104个,在常氧和低氧条件下孵育24 h。根据试剂盒说明书检测每个细胞培养上清液中的乳酸浓度。
1.2.3 线粒体DNA拷贝数分析
使用酚-氯仿法提取细胞样本的DNA,测定DNA浓度。将待测样品稀释为5 ng/μL。总反应体系:DNA 2 μL,2×SYBR Green 7.5 μL,上下游引物(10 μM)共3 μL,ddH2O 2.5 μL。使用实时荧光定量PCR仪进行待测样品扩增,扩增条件:预变性(95℃)10 min,变性(95℃)15 s,退火(60℃)延伸34 s,共40个循环。核DNA RPL13A基因上游引物:5′- CGCCCTACGACAAGAAAAAG-3′,下游引物:5′- CCGTAGCCTCATGAGCTGTT-3′;线粒体DNA mtCO1基因上游引物:5′-CAGGAGTAGGAGAGAGGGAGGTAAG-3′,下游引物:5′-TACCCATCATAATCGGAGGCTTTGG-3′。采用2-ΔΔCT 法分析mtCO1基因水平。
1.2.4 自噬相关蛋白LC3B、HSP60、TOM20表达检测
收集低氧组和常氧组BEAS-2B、A549细胞,分别加入200 μL RIPA细胞裂解液经超声波细胞粉碎仪破碎、离心后提取蛋白,并用BCA法测定蛋白浓度,并将等量的蛋白(15 μg)加载到SDS-PAGE凝胶中进行电泳分离蛋白后,转移到PVDF膜上,用10%的脱脂奶粉封闭150 min后,用PBST缓冲液洗去膜表面多余的脱脂奶粉,按照不同目的蛋白大小将膜切开并放入封口膜中,分别加入3 mL一抗孵育(4℃,过夜,β-Actin 1:1000,LC3B 1:1000,HSP60 1:1000,TOM20 1:4000)。用 PBST缓冲液洗膜后,加入3 mL山羊抗兔或山羊抗鼠 IgG(1:10000)孵育(室温)1 h后,用PBST缓冲液洗膜30 min,加入适量超敏ECL化学发光试剂,放入暗匣中曝光、显影。应用Image J软件分析蛋白条带的灰度值,以β-Actin蛋白为内参照,对以上目的蛋白进行灰度值分析,目的蛋白相对表达量=目的蛋白A值/内参照A值。
1.2.5 线粒体及溶酶体荧光强度共定位情况检测
将BEAS-2B、A549细胞接种于96孔板中,每种细胞接种12个孔(每孔接种100 μL细胞悬液,含4000个细胞),分设0、5、10 mM乳酸组;细胞贴壁后加乳酸,24 h后,弃原有培养基,加入100 μL含Mito tracker(线粒体染料)、CytoPainter LysoBlue(溶酶体染料)的培养基,Mito tracker的工作浓度为100 nM,CytoPainter LysoBlue的工作浓度为50 nM。染色(37℃,30 min)后弃含染料的培养基,加入100 μL PBS洗涤一次,再加入100 μL PBS,通过Cytation5酶标仪检测线粒体和溶酶体平均荧光强度及共定位情况。Mito tracker、CytoPainter LysoBlue检测分别使用Texas Red和DAPI通道。
1.2.6 统计学分析
2 结果
2.1 低氧条件对BEAS-2B、A549细胞内乳酸生成的影响
在常氧和低氧(1% O2)条件下培养BEAS-2B、A549细胞24 h,检测其培养上清液中的乳酸含量。结果显示,低氧培养条件下,BEAS-2B、A549细胞内的乳酸生成明显升高,差异有统计学意义(表1)。

表1 常氧和低氧条件下培养的BEAS-2B、A549细胞内的乳酸生成情况
2.2 低氧和乳酸对线粒体DNA拷贝数的影响
相比于常氧培养组,低氧条件下A549细胞的线粒体DNA拷贝数增加,BEAS-2B细胞线粒体DNA拷贝数减少,差异有统计学意义(P<0.0001)。由于低氧能使细胞内的乳酸含量升高,故使用外源乳酸干预A549细胞分析乳酸对线粒体DNA拷贝数的影响。结果显示,在常氧和低氧培养条件下,随着乳酸浓度的进一步增加,会引起A549细胞线粒体DNA拷贝数减少,BEAS-2B细胞线粒体DNA拷贝数增加。以上结果提示,外源乳酸干预能改变细胞线粒体DNA拷贝数,且对BEAS-2B、A549细胞的影响程度不同(表2)。
表2 常氧和低氧条件下经不同乳酸浓度处理的BEAS-2B、A549细胞线粒体拷贝数
2.3 外源乳酸对线粒体自噬的影响
前述结果显示,外源乳酸干预的BEAS-2B、A549细胞线粒体拷贝数发生改变,在此基础上分析乳酸对线粒体自噬的影响,以探索线粒体DNA拷贝数变化的原因。结果显示,与对照组相比,在常氧和低氧培养条件下用乳酸干预的BEAS-2B、A549细胞中蛋白LC3B-Ⅱ/Ⅰ比值增加(图1、2和表3),提示经乳酸干预后细胞自噬功能增强。进一步分析线粒体热休克蛋白(HSP60)和线粒体外膜蛋白(TOM20)的蛋白表达水平发现,经乳酸干预后,A549细胞中的HSP60、TOM20蛋白表达水平均降低(图1),可能是线粒体自噬增强引起细胞内线粒体含量减少所致;而在常氧和低氧条件下,乳酸干预的BEAS-2B细胞中的HSP60、TOM20蛋白表达水平未见明显改变(图2)。
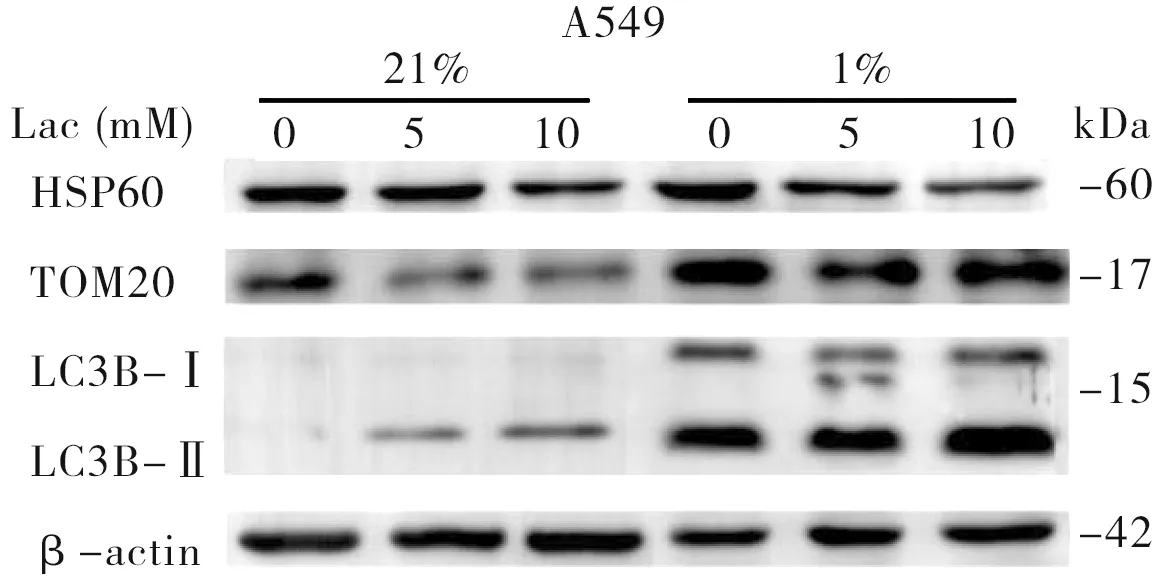
图1 在常氧和低氧条件下经不同浓度乳酸处理后的A549细胞LC3B、HSP60、TOM20蛋白表达水平
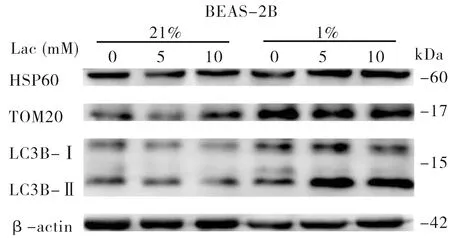
图2 在常氧和低氧条件下经不同浓度乳酸处理后的BEAS-2B细胞LC3B、HSP60、TOM20蛋白表达水平
表3 在常氧和低氧条件下用不同浓度乳酸处理BEAS-2B、A549细胞的LC3B蛋白表达量灰度值
2.4 外源乳酸对线粒体和溶酶体共定位信号的影响
由于线粒体自噬需要依赖溶酶体完成,故用线粒体和溶酶体荧光染料标记活细胞观察线粒体和溶酶体荧光信号共定位信号情况。分析结果显示,在常氧和低氧条件下,和对照组相比,乳酸干预后的A549细胞中线粒体和溶酶体共定位信号增多(图3、4和表4)。结合乳酸干预后的A549细胞中线粒体DNA拷贝数减少,线粒体HSP60和线粒体TOM 20蛋白表达水平下降,提示,外源乳酸可能引起A549细胞线粒体自噬增强。

图3 在常氧条件下经不同浓度乳酸处理后的A549细胞线粒体和溶酶体共定位信号

图4 在低氧条件下经不同浓度乳酸处理后的A549细胞线粒体和溶酶体共定位信号
表4 常氧和低氧条件下经不同乳酸浓度处理后的A549细胞线粒体和溶酶体共定位荧光强度值
3 讨论
近年来,免疫治疗的兴起为非小细胞肺癌的治疗带来了一线生机。机体免疫系统负责监控和杀伤肿瘤细胞,为了躲避免疫攻击,肿瘤细胞常常增强一些关键的免疫抑制蛋白的表达,以程序性死亡分子1(programmed cell death protein1,PD1)及其配体(programmed cell death protein 1 ligand 1,PD-L1)最为常见,这些分子能抑制免疫细胞的激活,进而减弱免疫应答。免疫治疗则是通过上述免疫抑制蛋白的抑制剂来激活免疫效应细胞,以达到杀伤肿瘤细胞的目的[1]。而被肿瘤浸润的淋巴细胞则是免疫治疗中被激活的主要效应细胞[2]。乳酸作为肿瘤微环境(TME)中的主要代谢物之一,它可以调节先天性免疫细胞和适应性免疫细胞的代谢,抑制CD+8T细胞和自然杀伤细胞、树突状细胞的活化和增殖。越来越多的研究表明,从癌细胞或间质细胞流出的质子偶联乳酸在保持酸性表型方面发挥了关键作用,并通过调节TME加速肿瘤进展,包括细胞侵袭、血管生成、转移发展和逃避免疫监视[3]。在肺癌模型中也发现了乳酸化的组蛋白,提示乳酸在肺癌细胞中很可能也通过组蛋白乳酸化修饰调控肺癌细胞和微环境组分细胞的基因表达[4]。可见,深入了解肺癌的免疫微环境特征是理解非小细胞肺癌肿瘤免疫调控机制及寻找新的干预靶点的关键。故本研究通过探求由低氧和乳酸构成的肿瘤微环境对肺癌细胞线粒体自噬的影响,寻找免疫靶点与疾病的相关性。
线粒体功能障碍所导致的氧化应激是癌症的主要驱动因素之一[5]。线粒体是所有真核细胞的 “动力工厂”,其主要功能是通过氧化磷酸化为细胞提供三磷酸腺苷(ATP),同时,还参与细胞代谢、增殖、衰老及程序性死亡等过程[6]。对于正常细胞来说,主要依赖线粒体氧化磷酸化来获取生命活动所需的ATP,而在肿瘤细胞中却普遍存在能量代谢方式的改变情况,被称为“有氧糖酵解”,即在氧气充足的条件下,肿瘤细胞仍然采用糖酵解的方式产生ATP[7]。线粒体是高等动物细胞核外唯一含有DNA的细胞器。mtDNA的功能实现不但与其结构完整性有关,还与其拷贝数有关。线粒体DNA拷贝数代表了每个细胞的线粒体数目及线粒体基因组数目,是衡量线粒体功能的指标之一[8]。本研究结果提示,人非小细胞肺癌细胞(A549)在无外源乳酸干预的情况下,低氧组相较于常氧组,线粒体DNA拷贝数有所增加,但在常氧和低氧条件下随着乳酸浓度的增加,线粒体拷贝数减少,说明低氧环境和乳酸对A549细胞的影响是不完全一致的,可能在于低氧环境不单增加乳酸浓度还会产生其他影响,如HIF信号通路的激活等[9,10]。
自噬是一种高度保守的自我消化和分解代谢过程,受损的蛋白质和细胞器聚集在自噬体内,然后通过与溶酶体融合来降解,以维持细胞的新陈代谢和稳定[11]。肿瘤细胞发生自噬的诱因源于营养和氧气供应不足、细胞毒剂的毒害等。[12-14]。大量研究证实,自噬对肿瘤的发生有积极的调控作用。微管相关蛋白轻链3B(LC3B)是最常用的自噬相关标记[15],是癌症中最常见的LC3亚型,在自噬过程中,LC3-Ⅰ通过类泛素系统的脂化作用转移到LC3-Ⅱ中。LC3-Ⅱ的含量与自噬水平相关,因此可作为自噬水平的标志。热休克蛋白60是一种重要的分子伴侣蛋白[16],位于2号染色体上,由核基因HSPD1[17]编码,主要分布在线粒体中,在辅助伴侣蛋白HSP10的帮助下发挥作用,纠正新生蛋白质的折叠,恢复错误折叠的蛋白质的结构,并维持线粒体蛋白质的稳定状态[18],是维持线粒体呼吸链的完整性和功能的正常性以及细胞生存所必需的过程[19]。故本研究首先检测自噬蛋白LC3BⅡ/Ⅰ比值,得出在常氧或低氧条件下,经外源乳酸干预后,非小细胞肺癌细胞(A549)较正常肺上皮细胞(BEAS-2B)自噬增加的结果,而后通过检测HSP60和线粒体TOM20的表达水平,分析线粒体与溶酶体荧光共定位信号的情况,进一步验证了乳酸与非小细胞肺癌细胞(A549)线粒体自噬的发生有关。
综上所述,在常氧或低氧条件下,乳酸浓度的上调可以通过增加线粒体自噬和减少线粒体拷贝数对人非小细胞肺癌细胞线粒体功能产生影响。在肿瘤早期,细胞相对处于常氧环境下,自噬可通过清除损伤的大分子或ROS产物抑制肿瘤的进展。而在肿瘤晚期,肿瘤细胞处于低氧条件下,自噬通过维持肿瘤细胞的生存促进肿瘤的发生发展。有研究证实,线粒体自噬可促进肿瘤的发生。在肿瘤转化过程中,致癌KRas细胞会诱发线粒体自噬的发生。在低氧条件下,肿瘤细胞发生糖酵解,并产生大量乳酸。ATP能量减少致细胞发生凋亡。然而,线粒体自噬可以克服这种能量的缺乏[20,21]。因此,研究乳酸在肿瘤细胞中的作用机制,有助于通过免疫治疗攻克非小细胞肺癌的研究;进一步研究在低氧状态下,乳酸对肿瘤细胞增殖、迁徙和侵袭的影响,可为全面攻克非小细胞肺癌发挥重要作用。
